



چگونه مقاله بنویسیم؟
- چگونه مقاله منتشر کنیم؟
- پزشکی مبتنی بر شواهد
- مرور سیستماتیک جناب آقای دکتر مجد زاده و همکاران
- داوری و نقد مقاله
- جستجوی منابع الکترونیک سطح 1
- جستجوی منابع الکترونیک سطح 2
- جستجوی منابع الکترونیک سطح 3
- EndNote
- Reference Manager
- مروز سیستماتیک جناب آقایان دکتر پیام کبیری و دکتر علی میرزازاده
- مرور سیستماتیک سرکار خانم ها دکتر نجات و دکتر غلامی و جناب آقای دکتر استوار
عفونت توام ناشي از ليشمانيا و HIV (Leishmania / HIV Co-infection)
ليشمانيوز
سالانه حدود 2-5/1 ميليون مورد ليشمانيوز پوستي و 500000 مورد ليشمانيوز احشائي در سطح جهان، رخ مي دهد. ليشمانيوز احشائي، شديدترين چهرة باليني ليشمانيوز است كه به دو شكل اپيدميولوژيك، بروز مي نمايد :
شكل زئونوتيك (Zoonotic) در حوزه مديترانه كه سگ به عنوان منبع اصلي عفونت براي حشرة ناقل، مطرح مي باشد
شكل انساني (Anthroponotic) در شرق آفريقا، بنگلادش، هند و نپال كه از طريق پشه خاكي سندفلاي، از انسان به انسان منتقل مي شود.
در افرادي كه دچار نقص ايمني نمي باشند عفونت حاصله در اغلب موارد به صورت بدون علامت يا تحت باليني، باقي مي ماند و در موارد با علامت باليني، معمولا به دنبال پشت سر گذاشتن دوره كمون چندهفته تا چندماهه، علائمي نظير: تب، كاهش وزن شديد، هپاتواسپلنومگالي و پان سيتوپني، عارض مي گردد و در صورت عدم درمان، ممكن است به مرگ بيماران منجر شود0
عفونت همزمان ليشمانيا/HIV
شكل باليني اصلي اين بيماري شامل وقوع همزمان ليشمانيوز احشائي و HIV/AIDS است كه در سال هاي اخير، بويژه در جنوب غربي اروپا باعث طغيان هاي متعددي گرديده است ولي ندرتا ممكن است با وقوع توأم ليشمانيوز پوستي و HIV/AIDS نيز مواجه شويم. نوپديدي ليشمانيا/HIV حاصل گسترش جهانگيري ايدز در مناطق روستائي و توسعة ليشمانيوز احشائي به حاشيه شهرها است. اين نوپديدي باعث تاثير عظيمي بر جنبه هاي باليني، تشخيصي، درماني و اپيدميولوژيك ليشمانيوز، گرديده است.
هرچند اين نوپديدي در بيش از 33 كشور، رخ داده است ولي بيشترين موارد آن در جنوب غربي اروپا و بخصوص در فرانسه، ايتاليا، پرتغال و اسپانيا حادث گرديده است (نقشه 4).
توزيع سني بيماران
طي سال هاي اخير، در بسياري از كشورهاي اروپائي، تغييرات اپيدميولوژيك عظيمي در الگوي ليشمانيوز احشائي، حاصل شده است. به طوري كه ليشمانيوز احشائي كه معمولا در بين كودكان حادث مي گرديد امروزه بيشتر در بالغين، رخ مي دهد و حدود 9/76% موارد در سنين 50-31 سالگي مي باشند كه منطبق بر محدوده سني معتادان تزريقي مبتلا به HIV/AIDS است (نمودار 25 و 26).
عفونت همزمان HIV و گونه هاي ليشمانيا به نحو شايعي به صورت ليشمانيوز احشائي، تظاهر مي نمايد.
وضعيت ايمونولوژيك بيماران
ابتلاء به ايدز، خطر بروز ليشمانيوز احشائي را به ميزان 1000-100 برابر، افزايش مي دهد و علاوه بر آن شدت ليشمانيوز احشائي در زمينه HIV/AIDS بيشتر از كساني است كه مبتلا به ايدز نيستند و جالب توجه است كه ليشمانيوز نيز اثرات متقابلي بر سير HIV/AIDS اِعمال مي كند. به طوري كه بر ميزان تكثير ويروس و شدت نقص ايمني، مي افزايد و در نتيجه، اغلب بيماران، داراي لنفوسيت هاي CD4+T كمتر از 200 مي باشند و طبيعي است كه تحت چنين شرايطي ساير عفونت هاي فرصت طلب نظير توبركولوز، كانديدياز، پنوموني ناشي از پنوموسيستيس كاريني . . . و توكسوپلاسموز نيز با شيوع بيشتري عارض مي شوند.
تشخيص باليني
تشخيص ليشمانيوز احشائي در مبتلايان به ليشمانيا/HIV، مشكل مي باشد. زيرا تظاهرات باليني شايع بيماري نظير تب، كاهش وزن و ارگانومگالي، هميشه وجود نداشته و يا تحت تاثير علائم مشترك ناشي از ساير عفونت هاي فرصت طلب، قرار مي گيرد. با اين وجود براساس مطالعه اي كه در بيماران اروپائي صورت گرفته است اين بيماران در اغلب موارد دچار علائم اصلي ليشمانيوز احشائي بوده اند. شايان ذكر است كه در بيماران مبتلا كه HIV/AIDS كه ضمنا دچار تب، ارگانومگالي و آنمي نيز مي باشند بايد با ظنّ باليني قوي به كالاآزار نيز فكر كرد و در مورد سابقه سكونت يا مسافرت به مناطق آندميك بيماري، تحقيق نمود.
|
|

نقشه 4 ـ انتشار جغرافيائي ليشمانيا / HIV
|
|

نمودار 25 ـ توزيع سني مبتلايان به ليشمانيا/HIV و معتادان تزريقي در كشورهاي اروپائي
|
|

نمودار 26 ـ توزيع بيماران مبتلا به ليشمانيا/HIV برحسب زمينه هاي مختلف
|
|

نمودار 27 ـ ميزان حساسيت تست هاي سرولوژيك كالاآزار در مبتلايان به ليشمانيا/HIV
تشخيص سرولوژيك
پاسخ منفي ايمني هومورال در 6/42% مبتلايان به ليشمانيا/HIV جلب توجه مي كند (نمودار 27) و از آنجا كه بر حسب نوع آزمون مورد استفاده ممكن است پاسخ هاي متفاوتي دريافت شود توصيه شده است همواره از تعداد 2 يا چند تست سرولوژيك، استفاده شود (نمودار 28).
|
|

نمودار 28 - ميزان مثبت شدن تست هاي سرولوژيك كالاآزار برحسب نوع آزمون
تشخيص انگل شناسي
تشخيص پارازيتولوژيك به منطور كشف انگل ليشمانيا در ارگان هاي بدن، از ارزش والائي برخوردار است. آسپيراسيون مغز استخوان مخصوصا زماني كه به طور مكرر در طي روزهاي آغازين بيماري، تهيه شود يكي از حساسترين روش هاي تشخيصي به حساب مي ايد و در 5/93% موارد، نتايج مثبتي به بار مي آورد. ضمنا در صورت شروع درمان يا عود بيماري به منظور افزايش حساسيت تشخيصي، مي توان از كشت مغز استخوان، استفاده نمود. همچنين با توجه به تراكم انگل در خون محيطي از آزمون Buffy-coat نيز مي توان استفاده كرد.
گرفتاري سيستم ادراري تناسلي
همة بيماري هاي مقاربتي ممكن است باعث تسهيل انتقال HIV بشوند. از طرفي در افراد مبتلا به عفونت ناشي از HIV بيماري سيفيليس و عفونت ناشي از HPV با سرعت بيشتري به مراحل پيشرفته، سير نموده و با احتمال بيشتري باعث ايجاد كانسر دهانة رحم مي گردد. سيفيليس اوّليه يا ثانويّه معمولا همراه با مثبت شدن آزمون هاي سرمي بوده و با سرعت بيشتري به سمت نروسيفيليس، به پيش مي رود.
آلگوريتم 1 ـ مراحل تشيخص HIV/AIDS
|
|

پيشگيري اوّليه و ثانويه عفونت هاي فرصت طلب در زمينه HIV/AIDS
آنسفاليت توكسوپلاسمائي
پيشگيري از تماس ها
افراد مبتلا به عفونت ناشي از HIV بايد هرچه سريعتر از نظر وجود IgG ضد توكسوپلاسما به منظور كشف عفونت نهفته توكسوپلاسمائي مورد بررسي قرار گيرند و كليّه افراد مبتلا به عفونت ناشي از HIV و ازجمله افرادي كه فاقد آنتي بادي IgG ضد توكسوپلاسما هستند بايد مورد مشاوره قرار گيرند و آموزش هاي لازم در زمينه راه هاي پيشگيري از بروز عفونت توكسوپلاسمائي به آنان داده شود.
به اين بيماران بايد توصيه شود از خوردن مواد گوشتي كه به طور كامل پخته نشده است اكيدا خودداري كنند. لازم به تاكيد است كه حرارت داخل گوشت گوسفند، گوساله و خوك بايد به حدود 175-170 درجه سانتيگراد برسد و با توجه به اينكه تحت چنين شرايطي رنگ صورتي وسط گوشت از بين خواهد رفت لازم است اين نكته عملي مهم به افراد HIV مثبت، تعليم داده شود. همچنين به آنها آموخته شود كه بعد از تماس با گوشت خام و خاك، دستان خود را بشويند و از خوردن ميوه ها و سبزي هاي خام نَشسته نيز اكيدا پرهيز نمايند و اگر در منزل، گربه نگهداري مي كنند تخته كف لانه گربه را روزي يك بار به وسيله يك نفر HIV منفي غيرحامله، تعويض كنند و ضمن جلوگيري از خروج اينگونه گربه ها از منزل مانع تماس آنها با گربه هاي ولگرد شوند و همواره آنها را با غذاهاي پخته شده و يا غذاهاي خشك آماده، تغذيه نمايند و به منظور سهولت كار و معافيت از خدمت رساني افراد HIV مثبت به گربه ها احتياط واجب آنست كه توصيه شود از نگهداري گربه در منازل، خودداري كنند.
پيشگيري اوّليه (پيشگيري از بروز بيماري)
افرادي كه از نظر آزمون سرمي توكسوپلاسما مثبت هستند و لفنوسيت هاي CD4+ T آنها كمتر از 100 مي باشد بايد تحت پوشش پيشگيرنده داروهاي موثر بر آنسفاليت توكسوپلاسمائي قرار گيرند. لازم به يادآوري است كه در صورتي كه اين افراد تحت پوشش پروفيلاكسي با روزانه 2 قرص معمولي كوتريموكسازول به منظور پيشگيري از PCP هستند همين اقدام باعث پيشگيري از بروز آنسفاليت توكسوپلاسمائي نيز خواهد شد. ضمنا آتوواكن به تنهائي يا همراه با پريمتامين نيز موثر واقع مي گردد ولي تك درماني با داروهائي نظير داپسون، پريمتامين، آزيترومايسين يا كلاريترومايسين، توصيه نمي شود و افشانه پنتاميدين نيز موثر نمي باشد.
افرادي كه از نظر آزمون هاي سرمي توكسوپلاسما منفي هستند و تحت پوشش داروهاي پيشگيرنده PCP موثر بر آنسفاليت توكسوپلاسمائي نمي باشند بايد زماني كه تعداد لنفوسيت هاي CD4+ T آنها به كمتر از 100 رسيد مجددا از نظر عفونت توكسوپلاسمائي، بررسي شوند و به هنگام بروز تغييرات سرمي از منفي به مثبت، تحت پوشش پروفيلاكسي قرار گيرند.
زمان قطع پروفيلاكسي اوّليه
جوانان و بالغيني كه تحت پوشش درماني با HAART هستند در صورت پاسخ درماني و افزايش لنفوسيت هاي CD4+ T به بيش از 200 و تداوم آن به مدّت بيش از سه ماه مي توانند از ادامه پروفيلاكسي عليه توكسوپلاسموز، خودداري كنند.
پيشگيري ثانويّه (پيشگيري از عود)
بيماراني كه دوره درماني اوّليه ضد آنسفاليت توكسوپلاسمائي را پشت سر گذاشته اند بايد تا پايان عمر، تحت درمان سركوبگر نيز قرار گيرند مگر اينكه ايمني آنان تحت تاثير درمان با HAART به وضعيت طبيعي باز گردد.
در اين بيماران مصرف توام پريمتامين + سولفاديازين + ويتامين B6 به منظور درمان سركوبگر، از تاثير والائي برخوردار است و در صورت عدم تحمّل سولفاميدها، مي توان از پريمتامين + كليندامايسين، نيز استفاده نمود. البته بايد توجه داشت كه سولفاديازين + پريمتامين از بروز PCP نيز جلوگيري مي كند.
زمان قطع پروفيلاكسي ثانويّه
جوانان و بالغيني كه تحت پوشش پروفيلاكسي ثانويّه براي آنسفاليت توكسوپلاسمائي هستند مشروط بر اينكه درمان آنسفاليت توكسوپلاسمائي را با موفقيت پشت سر گذاشته، فاقد علائم باليني بوده و تعداد لنفوسيت هاي CD4+ T آنها به طور پايداري در محدوده بيش از 200 باقي بماند از نظر عود آنسفاليت توكسوپلاسمائي در معرض خطر بالائي نخواهند بود و تحت اين شرايط مي توان از ادامه درمان سركوبگر، خودداري نمايند.
شروع مجدد پيشگيري ثانويّه
در صورتي كه تعداد لنفوسيت هاي CD4+ T به كمتر ار 200 سلّول كاهش يابد بايد رژيم درماني سركوبگر را مجددا آغاز نمود.
وضعيت هاي خاص
كودكان
در صورتي كه كودكان، تحت پوشش پروفيلاكسي PCP با كوتريموكسازول باشند در مقابل آنسفاليت توكسوپلاسمائي نيز محافظت مي گردند و آتوواكن نيز در آنان موثر خواهد بود.
كودكان بيش از 12 ماهه اي كه تحت پوشش پروفيلاكسي PCP هستند و داروهائي غير از كوتريموكسازول يا آتوواكن دريافت مي نمايند بايد از نظر عفونت توكسوپلاسمائي نيز بررسي شوند. از طرفي توصيه شده است كودكاني كه دچار سركوب شديد سيستم ايمني هستند و آزمون سرمي آنها از نظر آنتي بادي ضد توكسوپلاسما مثبت است بايد عليه توكسوپلاسموز و PCP تحت پوشش پيشگيري داروئي، قرار گيرند و داپسون باضافه پريمتامين، دريافت كنند. همچنين كودكاني كه سابقه ابتلاء به توكسوپلاسموز را داشته اند به منظور پيشگيري از عود بيماري، لازم است تا پايان عمر، تحت پوشش پروفيلاكسي، قرار گيرند.
شايان ذكر است كه وضعيت كودكاني كه تحت پوشش پروفيلاكسي اوّليه يا ثانويّه بوده اند به خوبي مطالعه نشده است.
زنان باردار
كوتريموكسازول را به همان شيوه اي كه به منظور پروفيلاكسي PCP توصيه كرده اند براي پيشگيري توكسوپلاسموز نيز قابل توصيه است ولي از آنجا كه از يك طرف خطر بروز آنسفاليت توكسوپلاسمائي، در دوران بارداري در حد پائيني قرار دارد و از طرف ديگر خطر بالقوه پريمتامين در اين دوران به اثبات رسيده است، توصيه كرده اند تجويز رژيم هاي داروئي حاوي پريمتامين، به بعد از ختم بارداري، موكول گردد.
در موارد نادري زنان حامله HIV+ كه داراي شواهد سرولوژيك عفونت توكسوپلاسمائي قديمي بوده اند عفونت را به جنين خود منتقل كرده اند و لذا توصيه شده است زنان باردار HIV+ كه دچار عفونت اوّليه توكسوپلاسمائي يا بيماري توكسوپلاسموز و از جمله آنسفاليت توكسوپلاسمائي هستند به دقت مورد ارزيابي و درمان قرار گيرند و نوزادان آن ها نيز پس از تولّد از نظر توكسوپلاسموز مادرزادي، مورد بررسي قرار گيرند.
كريپتوسپوريديوز
پيشگيري از تماس ها
افراد مبتلا به عفونت ناشي از HIV بايد در مورد راه هاي مختلف انتقال كريپتوسپوريديوم، تحت آموزش قرار گيرند. اين راه ها عبارتند از تماس مستقيم با بالغين مبتلا، شيرخواران، حيوانات ، آب آشاميدني و غذاهاي آلوده. اين بيماران بايد از تماس با مدفوع انسان و حيوانات، اجتناب كنند و در صورت تماس، دستان خود را به دقت بشويند. همچنين بعد از تماس با حيوانات خانگي و پس از تماس با خاك (باغباني و . . .) نيز به شستشوي دستها بپردازند و از رفتارهاي جنسي كه منجر به تماس مدفوعي ـ دهاني مي شود نيز اكيدا اجتناب نمايند.
اين افراد نبايد از آب جويبارها و رودخانه ها بنوشند و نسبت به اين واقعيت كه بسياري از آب ها ممكن است آلوده به مدفوع انسان يا حيوانات و به تبع آن آلوده به كريپتوسپوريديوم هستند آگاهي كافي داشته باشند و حتي از شنا كردن در آب هاي مشكوك، بپرهيزند و در صورتي كه در اينگونه آب ها شنا مي كنند بايد از بلع آب به طور جدّي خودداري كنند. ضمنا بايد توجه داشته باشيم كه يخ هائي كه از آب آلوده به كريپتوسپوريديوم تهيه مي گردد نيز ممكن است منبع عفونت باشند و آب ميوه ها و بسياري از نوشيدني هاي غيرپاستوريزه ديگر نيز ممكن است آلوده باشند.
شايان ذكر است كه در صورت آلودگي آب آشاميدني به كريپتوسپوريديوم، جوشاندن آن به مدت يك دقيقه باعث از بين رفتن اووسيست هاي انگل مي گردد. در مجموع با توجه به اينكه به دلايل مختلفي ممكن است آب هاي آشاميدني، آلوده به اين انگل شوند توصيه شده است افراد HIV+ سعي كنند همواره از آب جوشيده استفاده نمايند. ضمنا افراد HIV+ بايد از مصرف صدف خوراكي خام نيز بپرهيزند زيرا اووسيست هاي كريپتوسپوريديوم ممكن است تا بيش از دو ماه در بدن صدف زنده بمانند.
در بيمارستان ها، رعايت احتياط هاي استاندارد، معمولا مانع انتقال عفونت از افراد بيمار توسط كاركنان مي شود ولي از آنجا كه ممكن است اووسيست ها از طريق وسايل نير انتقال يابند برخي از متخصصين، توصيه كرده اند افراد مبتلا به عفونت ناشي از HIV و مخصوصا افرادي كه دچار سركوب شديد ايمني هستند نبايد با مبتلايان به كريپتوسپوريديوز در يك اطاق بستري گردند.
پيشگيري از بروز و عود بيماري
بيماراني كه ريفابوتين يا كلاريترومايسين به منظور پروفيلاكسي MAC مصرف مي كنند ممكن است در مقابل كريپتوسپوريديوزيس نيز محافظت شوند. ولي با اين وجود هنوز دلايل كافي براي تجويز اين داروها به عنوان پروفيلاكسي كريپتوسپوريديوزيس وجود ندارد. همچنين در مورد پيشگيري از عود نيز رژيم داروئي شناخته شده اي وجود ندارد.
ميكروسپوريديوز
در مجموع، غير از توجهات كلّي و رعايت موازين بهداشتي و بخصوص شستشوي دست ها اقدام ديگري توصيه نشده است و رژيم داروئي پيشگيرنده موثري وجود ندارد.

کریپتوسپوریدیوم، برای اولین بار توسط Tyzzer در سال 1907 در مخاط معده موش آزمایشگاهی مشاهده شد. برای 70 سال خصوصیات این تک یاخته به طور کامل معلوم نبود. توجه به این انگل در دو دهه گذشته اقزایش یافته است. این انگل از آنجا که در بیماران نقص سیستم ایمنی، تمایل به ماندگاری دارد، موجب اسهال مزمن در این بیماران می شود و از این نظر مورد توجه محققان قرار گرفته است و مکانیسم پاتوژنز وپاتوفیزیولوژی آن تحت بررسی می باشد.
کریپتوسپوریدیوم، زئنوتیک است و افرادی که با دامها تماس دارند در خطر ابتلا به آن هستند. کودکان با سوء تغذیه نیز در معرض ابتلا به اسهال مزمن با این انگل هستند. عفونت، ماهها و حتي سالها در افراد با نقص اكتسابي سيستم ايمني و يا نقص مادرزادي آن باقي ميماند و حتي ميتواند به سمت كبد و صفرا و يا لولههاي پانكراس گسترده شود. کریپتوسپوریدیوم 25% موارد اسهال در دنیا را به خود اختصاص داده است و انگلی جهان شمول است. کریپتوسپوریدیوم، جزء فیلوم اپی کمپلکسا (دارای کمپلکس راسی )، کلاس اسپوروزوئیتها (دارای سیکل زندگی جنسی و غیر جنسی و اووسیست )، زیر کلاس Coccidiasina ( دارای موروگونی و گامتوگونی) تحت راسته Eimeriina، راسته کوکسیدیاهای حقیقی، از خانواده Cryptosporidae است.
تا سال 1955 این تک یاخته یک ارگانیسم کامنسال تصور می شد تا اینکه در این سال، عامل انتریت کشنده بوقلمون مشاهده شد. در سال 1971، اولین مورد کلنیکی آن را در گوساله های ایالات متحده آمریکا گزارش کردند. در دهه 1976 اولین اطلاعات در مورد پاتوژنز انسانی کریپتوسپوریدیوم توسط Nime ، گزارش شد.
95سال پس از پي بردن به بسياري از بيماريهاي رودهاي، كريپتوسپوريديوم انگلي كه مهرهداران را آلوده ميكند ، هنوز به صورت معمائي باقي مانده است.

كريپتوسپوريديوم ، انگلي با انتشار جهاني است و بيست گونه از آن در مهره داراني چون پستانداران، پرندگان، خزندگان و ماهيها گزارش شده است. از ميان گونههاي مختلف، كريپتوسپوريديوم پارووم انسان و ساير گونهها را اغلب آلوده ميكند. مشخصات مولكولي كريپتوسپوريديوم، باعث تفكيك گونه پارووم به دو بخش شده است، گروهي كه مربوط به انسان ميباشد و گروهي كه مربوط به اغلب جانوران است. مكانيسم واكنش كريپتوسپوريديوم با سلولهاي ميزبان هنوز مبهم باقي مانده است. تاكسونومي اين پارازيت مبارزهاي جدي را از بين بيولوژيستها و اپي دميولوژيستها در زمينه مولكولي برانگيخته است. مشكل، شناخت فنوتيپ در گونههاي ميزبانان مختلف ميباشد (گرچه اين تفاوتها آشكار هستند) و اين در حالي است كه كريپتوسپوريديوم از منابع مختلف بدست آورده ميشود. گوسفندان (با 7 گونه حيواني آن) و انسان با اسهالهاي شديد عفوني شدند. اين انتقال بين حيوان و انسان مشخص كرد كه انگل يك زئونوز است، كه باعث اسهال شديد شده است. بر پايه اين مدارك اوليه، نامگذاري گونه كريپتوسپوريديوم صورت گرفت و اين در حالي بود كه آنها هنوز به خوبي شناخته نشده بودند. كريپتوسپوريديوم پارووم مهمترين گونه ايجاد عفونت بادوام درپستانداران است. شمار دقيق گونههاي اضافه شده هنوز معلوم نيست. اما با استفاده از متدهاي مولكولي، دانش ما درباره تاكسونومي اين گونه افزايش يافته است.
21تفاوت عمومي تاكنون بين گونهها مشخص شده است و در حال حاضر حداقل 6 جنس از كريپتوسپوريديوم شناخته شدهاند.
هيچ ارگانيسمي مثل كريپتوسپوريديوم، ديواره سلولي را براي ايجاد موقعيت و جايگاه خودش تغيير نميدهد.نه مكانيسمهاي اين پروسه و نه مكانيسمهاي انگل به طور دقيق درك نشدهاند! اين ويژگي كريپتوسپوريديوم را به رازي براي شميوتراپي تبديل كرده است. كريپتوسپوريديوم مثل بقيه كوكسيدياها در خلال رشدش، خودش را درون سلول ميزبان مخفي ميكند. اين روند، مكانيسمي براي محافظت انگل از پاسخ سيستم ايمني ميزبان و از محيط روده است و اين در حالي است كه تغذيه و منبع انرژي انگل از ميزبان فراهم ميشود (مانند ساير كوكسيدياها).
كريپتوسپوريديوم، در درون سلولهاي ميزبان در يك حفره به نام حفره پارازيتيفوروس قرار ميگيرد و مجرايي از سيتوپلاسم ميزبان براي خود فراهم ميآورد تا واردانگل شود و آن را تغذيه كند. كريپتوسپوريديوم برخلاف هر كوكسيدياي ديگر، يك ساختمان منحصر به فرد به عنوان غشاء تغذيه كننده ارگانل دارد كه مستقيما بين سلول و سيتوپلاسم ميزبان گسترده ميشود. تصور ميشود كه (PVM) phorous vaculolar membrane فقط نقش محافظتي را براي آن دارد (در حالي كه غشاء منع تغذيه ارگانل است و جايگاهي براي برداشت انرژي از سلول ميزبان است). همچنين (PVM) نفوذپذيري انتخابي نسبت به بعضي از مواد معين از لومن روده دارد. اين حقيقي است كه PVM ميتواند عملكردهاي ديگري نيز براي انگل داشته باشد و اين در حالي است كه PVM معمولا از غشا سلول ميزبان مشتق ميشود.

مرگ داخل سلولي كريپتوسپوريديوم پارووم
عوامل ضد ميكروبي به منظور تاثير موثر بر ميكروارگانيسمهاي داخل سلولي بايد وارد سلول شوند و مكانيسم تا از بين بردن انگل ، داخل سلول ميزبان ادامه مييابد.
مكان مخصوص C.parvum زير غشاء سلول يا PVM يا در لومن روده است. اين انگل مستقيما به دومنهاي خارج سلولي دسترسي دارد.
براي تحقيق در باره دارويي كه بتواند مانع انتقال C.parvum شود، ما بايد از پارومومايسين استفاده كنيم. تنها عامل آنتي ميكروبيال كه به طور جزئي ولي سازگار ميتواند اثر بگذارد و از گسترش و رشد انگل در invitro , invivo جلوگيري مي كند. اين مطالعات نشان ميدهد كه، پارومومايسين و جنتيسين ،كه يك آمينوگلي كوزيد ديگر است، ميتوانند فعاليت رادر invitro مهار كنند.
غشاهايي كه از انگل و حفره پارازيتيوفوروس منشا گرفتهاند بيشك سهم بزرگي در ورود پارومومايسين دارند و امكان دارد پذيراي بقيه داروها هم باشند. اين مشاهدات ميتواند براي، تعيين هدف داروها مورد توجه باشد.
پاتوفيزيولوژي
مكانيسم بيماريزايي كه C.parvum ايجاد ميكند منجر به اسهال ، سوء جذب vitB12 ، D- گزيلوز و استئاتوره ميشود. اين مكانيسمها اولين واكنش انگل و ميزبان از حمله انگل به بافت رودهاي ميزبان است.
اولين واكنش بعد از ورود انگل به سلول ميزبان اين است كه، عامل تهاجم وحفره پارازيتيفوروس باهم يكي ميشوند كه شامل چندين ليگاند و گيرنده براي انگل و رسپتورهاي ميزبان است.
اين واكنش در اپي كمپلكسا مثل توكسوپلاسما همچنين در پلاسموديوم و امريا نيز مطالعه شده است.
مرحله زوئيت غيرتهاجمي شامل رافتيدي و ميكرونمها و گرانولوهاي دنس يا متراكم هستند كه همه جزء كمپكلس رأسي ميباشند.
در خلال اولين واكنش انگل – ميزبان، اين ارگانها به طور مداوم و پيدرپي پروتئينهاي اگزوسيتوز را ترشح ميكنند كه تهاجم را براي انگل مهيا ميكند و همچنين باعث شكلگيري واكوئل ميشود. بسياري از پروتئينهاي ميكرونمال، مولكولهاي چسبندگي دارند كه انگلهاي گروه اپي كمپلكسا را از صدمه مصون نگه ميدارند. به منظور افزايش شناخت كريپتوسپوريديوم به عنوان يك پاتوژن انساني بايد پروتئينها و كمپلكس راسي آن شناخته شود اين پروتئينها شامل:
CSL,GP900,P23/27,TRAPC1,GP15,CP15,GP60,CP47,gp40/45 and gp15/Cp17 ، میباشد.
از مطالعاتي كه در باره اين پروتئينها صورت گرفته است، اين گونه استنباط ميشود كه در بيماريزايي كريپتوسپوريديوم نقش دارند.
پروتئین های موثر در پاتوفیزیولوژی
CSL: circumsporozoite - like antigen
يك گلي كوزيلات با وزن 1300KD است كه بسيار گلي كوزيله شده است و با استفاده از آنتيبادي مونوكلونال كه به آن اپي توپهاي تكراري كربوهيدراتي متصل كرده بودند شناخته شد.
اين mAb دلالت به همان circumspao ziote-like Ag دارند. همچنين اين mAb عفونت را در مدل موشي كريپتوسپوريديوم در ivitro,invitro خنثي كرده است.
CLS، بر روي ميكرونماها و گرانولهاي متراكم كمپلكس راسي و روي سطح اسپوروزئيت قرار گرفته است.
مطالعات جديد نشان ميدهد كه CLS به يك رسپتور KD85 بر روي سلولهاي اپيتليال رودهاي متصل ميشود، هنوزساختار مولكولي اين pro مبهم است. اين بافتها ميتواند CSL را به عنوان هدفي براي درمان كريپتوسپوريديوم مطرح كند.
GP900 Glicoprotein 900
مثل CLS يك گلي كوپروتئين با وزن مولكولي بالاست كه بسيار گلي كوزيله شده است. در ميكرونماي اپي كمپلكسا سنتز مي شود و در سطح زوئيتهاي مهاجم و در خلال حركت آنها به بيرون ترشح ميشود. آناليز و تجزيه سكانسهاي آمينواسيد GP900 نشان ميدهد كه، يك پروتئين مولتي دومن است كه غني از سيستئين ميباشد و دومينهاي شبه موسيني دارد (يك دومن بين غشايي و دم سيتوپلاسمي).
GP900 در سمت Nترمينال، گلي كوزيله شده است. GP900 به سطح سلولهاي اپيتليال رودهاي متصل ميشود. اين مشاهدات نشان ميدهد كه GP900، واسطه تهاجم به سلولهاي ميزبان است ارتباط بين CLS,GP900 هنوز به درستي معلوم نيست.
Thrombos pondin-related adhesive protein(TRAPC1)
پروتئينهاي چسبندگي وابسته به ترومبوسپوندين، هومولوگي از پروتئينهاي ميكرونمال است كه شامل TRAP, CTRP, CS, Etp100 and MIC-2 ميباشند.
اين پروتئينها يك دومين ترومبوسپونديني دارند و به وسيله ترومبوسپوندين مولتي پل وابسته به موتيفها نشان داده شدهاند و به طور قطع براي درمان سلولهاي ميزبان اهميت دارند.
محصول سكانس آمينواسيدهاي TRAPC1 يك سكانس Nترمينالي است كه يك دومين پليسرين است، يك دومن ترومبوسپونديني نيز دارد، يك دومن بين غشايي و يك دم سيتوپلاسمي است. با استفاده ازآنتيباديهاي نوتركيب شده، TRAPC1 را لوكاليزه ميكنند، هنوز هيچ شواهدي از درگيري TRAPC1 در تهاجم انگل به سلولهاي ميزبان ديده نشده است.
gp40
تحقيقات بيشتر ما را به سمت gp40 ميبرد، يك گليكوپروتئين O-گلي كوزيله شده شبه موسيني ديگر، كه در مرحله تهاجم انگل به سلولهاي ميزبان، بر روي سطح منطقه راسي قرار دارد و از سطح پارازيت بيرون ريخته ميشود. آنتيباديهاي اختصاصي ضد gp40 عفونت را در invitro خنثي ميكنند. Gp40 در C.parvum، به طور اختصاصي به سلولهاي ميزبان باند ميشود تصور ميشود كه اين پروتئين در چسبندگي و تهاجم نقش داشته باشد. كلون كردن ژن gp40 در C.parvum. آناليز آمينواسيدهاي آن يك سيگنال پپتيد N- ترمينالي و يك دومين پلي سريني را نشان داد.
· CPgp40/15
يك ايمنودمن 15/17KD را كد ميكند كه در واكنش انگل ميزبان دخيل است و در مرحله تهاجم در سطح انگل قرار داد.
در حال حاضر gp15,gp40 با مرحله بعد از ترجمه پروكوسور گليكوپروتئيني، از هم مشتق ميشوند كه به وسيله Gp40/15 در مراحل خارج سلولي انگل كد ميشوند gp40، قطعه N ترمينال محلول و gp15، C- ترمينال پروكورسور gp40/15 هستند كه در خلال مراحل بعد از ترجمه به صورت واحد باقي ميمانند.
CP47
يك پروتئين غشايي است، كه به سطح سلولهاي اپيتليال رودهاي باند ميشود و در قسمت راسي اسپوروزئيت است. هنوز ژني كه CP47 را كد ميكند كلون نشده است.
با توجه به قرائن اينطور استنباط ميشود كه پروتئينهاي كه توضيح داده شد هم در درمان و هم در تهاجم انگل ميتوانند دخيل باشد.
كريپتوسپوريديوز در مبتلایان به ایدز
كريپتوسپوريديوز، يكي از عفونتهاي فرصت طلب شايع در ميان مبتلايان به AIDS است. شمارش سلولهاي لنفوسيتي T4 در در خلال رشد انگل و سيكل زندگي آن در اين افراد به كمتر از ml/150 ميرسد.
عفونت، ماهها و حتي سالها در افراد با نقص اكتسابي سيستم ايمني و يا نقص مادرزادي آن باقي ميماند و حتي ميتواند به سمت كبد و صفرا و يا لولههاي پانكراس گسترده شود كه باعث Choledochitis , cholecystitis , cholangio hepatitis يا pancreatitis ميشود. تلاش محققان برای یافتن درمان موثر و یا واکسن مخصوص كريپتوسپوريديوم ادامه دارد.
عدهای از محققان در اواسط دهه 1990 درهند، شیوع اسهال كريپتوسپوريدیایی را در مبتلایان به ایدز بررسی کردند. بررسیهای آنان نشان داد که: اسهال كريپتوسپوريدیایی در 7% تا 83% از بیماران با علائم و در 4/1% تا 57% بدون علائم است. درصدهای مشابهی نیز از شمال شرقی آمریکا گزارش سده است. در بررسی با متدهای پیشرفته ای مانند Molecular tools و در بررسیهای اپیدمیولوژیک معلوم شده است که مبتلایان به ایدز( بالغین و کودکان)، مستعد آلودگی با گونه های مختلف كريپتوسپوريدیوم هستند.البته شيوع اين بيماري در مبتلايان به ايدز خيلي بالا نيست (15%-5 در كشورهاي توسعه يافته).
فقدان راههاي مناسب درماني براي اين انگل آنرا به يك مشكل در ميان عفونتهاي فرصت طلب كه با ايدز ديده ميشوند، تبديل كرده است.
كريپتوسپوريديوز در كودكان با سوء تغذيه
چندين گزارش نشان ميدهد كه اسهال كريپتوسپوريديايي در كودكان با سوء تغذيه تمايل به ماندگاري دارد. در مطالعه ای محققان 1779 کودک با اسهال را بررسی کردند. 444 کودک، علت اسهالشان كريپتوسپوريدیوم پارووم بود. آنالیز PCR بر روی 444 نمونه استول این کودکان نشان می داد که 74% با تیپ 1 انسانی آلوده شده اند و 19% با تیپ 2 ، 6% آلودگیها نیز مخلوطی از هر دو تیپ بود، آنان در یافتند که در کودکان با سوء تغذیه اسهال پایدار شیوع بیشتری دارد (31%) و 22% از کودکان نیز اسهال حاد داشتند. كريپتوسپوريدیوز در کودکان ایرانی نیز بررسی شده است. تحقیقاتی در بین می 2003 تا اکتبر 2003 در 614 کودک با علائم و بدون علائم در کلنیک کودکان در کرمانشاه، صورت گرفت. پژوهشگران از هر 515 کودک با علائم یک نمونه استول گرفتند و از هر99 کودک بدون علائم نیز یک نمونه گرفتند. بعد از تهیه اسمیر از نمونه ها آنها را با متد ذیل نلسون اصلاح شده رنگ کردند.در 4/10 % از نمونه ها C.parvum یافتند. درصد آلودگی با C.parvum ، به طور مشخصی در کودکان با اسهال نمودار بود. آنها در 2/ 15% از کودکانی که در روستا زندگی می کردند و در 2/ 7 % از کودکانی که در مناطق شهری بودند C.parvum را یافتند.درصد آلودگی با C.parvum در کودکانی که با حیوانات تماس داشتند 5/8 % بود. این درصد در کودکانی که با حیوانات تماس ندارند به 2/ 6% می رسد. مشکل ایجاد اسهال پایدار، دهیدراتاسیون و سوء جذب در این کودکان است.
در خلال 15ماه مطالعه در بيمارستان مالگو در كامپالا، از 13556 نفري كه از بچهها اسهال داشتند، 200 كودك وضعيت حادي داشتند.
در ميان 1779 كودك بيمار 9/29% كه اسهال در آنها بيشتر از 14 روز طول كشيد، در گروه اسهال مزمن قرار گرفتند. از 1779 كودك با اسهال در حدود 40% آن دچار دهيدراتاسيون شده بودند.
مطالعات عدهاي از محققين نشان داده است كه: ارتباطي قوي، بين سن و ايجاد شدن اسهال وجود دارد. تقريبا 25% بچهها در کودکی براي اولين بار دچار عفونت ميشوند.
آپوپتوز
براي بررسي ارتباط بين اين انگل و آپپتوز، در ابتدا بهتر است پديده آپوپتوز را بررسي كنيم.
آپوپتوز تحت كنترل ژنتيك است. در هسته سلول اتفاق ميافتد و هسته سلول را آرام آرام جمع ميكند مهمترين ويژگي آپوپتوز اين است كه ديواره از بين نميرود و پروتئوليز، در داخل سلول آرام آرام اتفاق ميافتد.
نكروز، يك عارضه پاتولوژيك است.در نكروز سوراخ روي ديواره سلول ايجاد ميشود. سلول متورم ميشود و سرانجام محتويات سلول خارج ميشود.
بعد از سالها، دانشمندان دريافتند آپوپتوز در تك سلوليها هم اتفاق ميافتد. در اين روند، 2 مسير داريم: مسير داخلي و مسير خارجي.
در مسير داخلي، ميتوز نقش مهمي دارد. آنزيمهايي كه در مرگ سلول نقش دارند و در ميتوز هم مهمند، فعال ميشوند. هر نوع آسيبي به DNA منجر به خارج شدن سيتوكروم C از انگل يا متازوآ ميشود، سيتوكروم C روي آنزيمها اثر ميكند كه مهمترين اين آنزيمها، كاسپاز است. در اثر فعال شدن كاسپاز مسير به صورت آبشاري تا خرد شدن DNA و پروتئوليز پروتئينها پيش ميرود.در مسير خارج سلولي نيز فعال شدن كاسپاز رخ ميدهد.
كاسپاز، آنزيم اصلي در القاء مرگ سلولي است. بعدها هومولوگي به جاي كاسپاز به نام متاكاسپاز را شناسايي و معرفي كردند. نام كاسپاز از سيستئين آسپارتيك اسيد پروتئاز گرفته شده است.
القاء آپوپتوز در سلولهاي رودهاي به وسيله C.parvum
كريپتوسپوريديوم پارووم، عامل اسهال در ميزبان با نقص ايمني و در ميزبان با سيستم ايمني كار آمد است. سايت اول C.parvum اپيتليوم سلولهاي رودهاي است، علاوه بر اين مجاري هوايي، شكم يا كيسه صفراء نيز ميتواند عفوني شوند. مرحله تهاجم انگل اووسيست است كه معمولا 4 اسپوروزوئيت را دارد. به دنبال پاره شدن آن،اسپوروزئيت به روده حمله ميكند و از طريق قسمت راسي خود گسترش مييابند. C.parvum در 48ساعت اوليه در كشت سلول و در invitro سيكل غيرجنسي خود را از سر ميگذراند. C.parvum باعث كوتاه شدن ويليها، هايپرپلازي كريپتها، و تغيير دادن پروتئينهاي ستيواسكتال ميشود و جذب سديم را هم تغيير ميدهد.
همچنين باعث افزايش محصولات پروستاگلايدين ميشود و كموكاينهاي اپتليال را براي ترشح تحريك ميكند.
ناحيهاي راسي C.parvum باعث آزاد شدن آنزيم سيتوليكي-لاكتات دهيدروژناز ميشود. اما هنوز نقش دقيق اين فاكتور شناسايي نشده است.
آپوپتوز، يك روند برنامهريزي شده از مرگ سلول است كه ميتواند به وسيله عوامل محيطي يا سلولهاي مجاور و ديگر رخ دهد، كه با نكروز سلولهاي اپيتليال فرق ميكند.
آپپتوز ميتواند در خلال ابتلا به چندين عفونت مهاجم يا غير مهاجم در بيماريهاي رودهاي انسان رخ بدهد، مثل گونههاي سالمونلا و شيگلا، در E.coli و يا ويروس نقص سيستم ايمني تيپ1، و هليكوباكترپيلوري.
يافتههاي محققين نشان داد كه در حضور فعاليت فاكتو ر NF-κβ، آپوپپتوز سلولهاي اپيتليال روده محدود ميشود و زماني كه فعاليت NF-κβ بلوكه ميشود، ظرفيت آپوپتوز به طور بارزي افزايش مييابد. اينكه آمادگي سلولها در اين حالت براي آپتوپوز بيشتر شده بود جالب توجه بود. عدهاي ديگر از محققين دريافتند كه پاتوژنهايي مانند توكسو پلاسما گوندي و كلاميديا ترايكومايتيس، با سلولهاي آلوده به C.parvum رقابت ميكنند و روند آپوپتوز را كاهش ميدهد. محققين دريافتند كه آپپتوز با درجات متوسطي به وسيله C.parvum در سلول القاء ميشود كه اين روند به رشد سلولهاي اپيتليال نيز بستگي داشت و دريافتند كه آپوپتوز و نكروز ميتواند توسط سيستم ايمني ميزبان محدود شود و حذف سلولهاي آلوده شايد به نفع ميزبان و محدود شدن واكنشهاي التهابي ميزبان باشد.
عدهاي ديگر از محققين اين تحقيقات را بر روي مدل شي، نيز انجام دادند. يافته هاي آنان، نشان داد كه زماني كه C.parvum وارد سلول ميشود، اپي تليال ايلئوم كاهش مييابد. ويليها كوتاه و ضخيم ميشوند و آپپتوز كمي هم مشاهده شد. اين يافتهها نشان ميدهد كه آپوپتوز يكي از مشخصههاي عفونت با C.parvum است.
محققين براي اين پژوهش ،10موش 7 روزه انتخاب كردند و آنها را در معرض آلودگي با C.parvum قرار دارند. بعد از گذشت زمان cm1 از انتهاي ايلئوم هر كدام را برداشت كردند و آنرا با فرمالين 10% فيكس كردند و براي برش در پارافين قرار دادند. در بعضي از موشها دئودنوم و كلون را نيز برداشت كردند برشها را با رنگآميزي هماتوكسيلين- ائوزين رنگ آميزي كردند. برشها را زير يك ميكروسكوپ نوري مورد بررسي قرار گرفت، طول ويليهاي روده را به وسيله يك ميكرومتر چشمي در يك ميكروسكوپ نوري بدست آوردند. بعد از 7 روز تعداد اووسيستهاي C.parvum به حداكثر رسيده بود. گرچه وزن موشها بعد از 7 روز عوض نشده بود يا علائم اسهال شديد را نشان ندادند. رنگآميزي H&E گرانولهاي ائوزينوفيليكي كه رنگ را به خود جذب كرده بودند نشان ميداد و همچنين سلولهاي گابلت در تمام موشهاي آلوده كاهش يافتهها بودند. يافتهای آنان نشان داد كه عفونت با C.parvum ميتواند به تغيير ساختار اپيتليوم ايلئوم و تغيير عناصر ساختار سلولي منجر بشود. اين تغييرات منجر به تغيير عملكرد روده و همچنين سوء جذب vitB12 نيز ميشود.
همچنين گزارش شده است كه پروتئين نوتركيب TAT در HIV ميتواند آپپتوز را به وسيله C.parvum افزايش بدهد.
با توجه به تمام يافتهها مكانيسم دقيق بيماريزايي درC.parvum مبهم باقي مانده است.
پاسخ سیستم ایمنی انسان به C.parvum
پاسخ سیستم ایمنی به انگل ذاتی واکتسابی است . فعالیت NFκβ منجر به آزاد شدن کموکاینها می شود. در لاین سلولهای روده ای در invitro آزاد شدن CXCL-8 در خلال عفونت با C.parvum مشاهده شده است. در مطالعه ای در برزیل محققان دریافتند که در استول کودکان بیشترIL-8 یافت می شود. در آنالیز بافت روده مبتلایان به ایدز CXCL-8 یافت نشده است. CCL5 نیز بعد از عفونت، در روده حضور می یابد. میزان این کموکاین در افراد با سیستم ایمنی کار آمد در مقایسه با بیماران نقص سیستم ایمنی تفاوتی ندارد.
در ابتداء کموکاینهایی مانند CXCL-10 و شاید CXCL-8 به عنوان کموتاکسی برای گرانولوسیتها عمل میکنند. فعالیت IL-15 نیز در ابتدای عفونت مورد توجه است. نقش کلیدی را Tcell CD4 و γIFN ایفا می کند. پاسخ التهابی بدن به انگل، موجب اسهال می شود و شاید TNF-β و IL-10 نقش ضد التهابی داشته باشند و باعث بروز IgA در مخاط روده می شوند، از این طریق از عفونت مجدد نیز جلوگیری می شود.
برگرفته از سایت:
http://parasitology.blogfa.com/post-30.aspx


|
|
|
A course on parasitology in English by Fred R. Opperdoes
Research Unit for Tropical Diseases,"Christian de Duve" Institute of Cellular Pathology and Laboratory of Biochemistry, Université Catholique de Louvain, Avenue Hippocrate 74-75, B-1200 Brussels, Belgium
Table of contents
-
Definitions and concepts
-
-
-
-
-
-
Novel drugs
-
-
Theileriosis
-
Schistosomiasis
-
Lymphatic filariasis
-
Onchocerciasis or river blindness
|
لیشمانیازیس
 |
|
لیشانیازیس بیماری مشترک انسان وحیوان است که میزبان مخزن بین حیوانات وحشی است. حیوانات اهلی به عنوان دومین میزبان مخزن به شمار می آیند.این انگل بصورت اماستیگوت درمهره داران وبصورت پروماستیگوت درحشرات دیده میشود. کالا آزار مشاهده نمودند. است کینتوپلاستیدها متعلق به شاخه او گلنوزوا که تک یاخته او گلنوئیدهم جزء همین شاخه است . لیشمانیا دارای دو مرحله تکثیر است ( Digenetic ) یا چند میزبانه (Hetero xenous ) است که بعنوان انگل در حشرات و مهره داران عمل می کنند میزبان های مهره دار شامل پستانداران و مارمولک هاست میزبان های پستاندار آن انسان ، سگ و بعضی جوندگان است . است . گونه های لیشمانیایی عهد جدید و عهد قدیم شناخته شده اند و در هر دو بیماری می تواند جلدی یا احشایی باشد . حشره های ناقل پشه خاکی از جنس فلوبوتوموس phlebotomus و لوتزومیا ( Lutzomyia ) است که در آنها انگل بصورت پروماستیگوت در لوله گوارش زندگی می کنند که در هنگام خونخواری از بخش پیشین لوله گوارش (foregut ) مسدود شده توسط انگل به زخم وارد میشود. دارد
های پستاندار بلعیده می شود. و تبدیل به آماستیگوت ( به اندازه ۴-۲ نانومتر ) می شود انگل در واقع درون فاگولیزوزوم ماکروفاژ زندگی میکنند آنها تقسیم شده و پس از ترکاندن سلول وارد سلولهای دیگر می شود در محل ابتدا تولید یک پاپول کوچک می کند. به خود بهبود می یابد و موسوم به زخم شرقی ( سالک ) می باشد در حالیکه انگل لیشمانیا دونوانی بداخل سلول های پوشینه ای سیستم تورینه – پوشینه ای و گلبول های سفید با هسته چند شکلی خون ، طحال ، غدد لنفاوی ، مغز استخوان و در نهایت ایجاد لیشمانیازیس احشایی می کند که در صورت عدم درمان ممکن است کشنده باشد. می کند که می تواند ریشه ( متاستاز) به بینی و دهان بدهد و باعث بد ریختی و بیماری غیر قابل درمانی را بنماید.
گوارش شده و در بخش میانی لوله گوارش ( Mid gut ) تکثیر یافته و تبدیل به پروماستیگوت شده به سرعت موجب انسدادلوله گوارش شده و واردمری میشود.
معمولا اماستیگوت ها در داخل سلول ها کشت یافته رشد داده می شوند اما می تواند در خارج سلولی در ۳۷ درجه در شرایط خاص رشد داده شود.
های لیشمانیا و هم واکنش ایمنی میزبان است عفونت لیشمانیا دونووانی با دوره کمون ۴-۲ ماه از بدون نشانه تا بروز بیماری کامل فرق می کند. یافته و در نهایت پس از ۳-۲ سال موجب مرگ می شود در بعضی از موارد که بیماری بصورت حاد است با تب بالا و لرز و در نهایت در ۱۲-۶ ماه موجب مرگ میگردد. علت مرگ معمولا در اثر عفونت های ثانویه است بعضی از عفونت ها هم خود بخود بهبود ی می یایند.
هند توسط ویلیام لیشمان کشف شد. ( بیماری در هند موسوم با کالاآزار یا تب دام دام است ) لیشمان این موضوع را در سال ۱۹۰۳ منتشر کرد و همزمان با او چارلز دونوان نیز موفق به کشف این انگل شد در بعضی از مناطق در ۱۰-۵ درصد از موارد درمان شده آشفتگی پوستی ایجاد می شود که موسوم به تظاهر لیشمانیایی جلدی پس از کالا آزار است ( PKDL)
گونه لیشمانیا رامعرفی نمودند تروپیکا(جلدی ) برازیلنیس ( جلدی مخاطی ) و دونووانی ( احشایی ) بهر حال طبقه بندی متداول خیلی پیچیده تر و باز تاب پیچیدگی نشانه های است بعنوان مثال انگل های کالا آزار هندی و مدیترانه لیشمانیا دونووانی و لیشمانیا اینفانتوم. ▪ لیشمانیازیس جلدی عهد قدیم و لیشمانیاماژور بعد از گزش پشه خاکی (فلوبوتوموس ) آلوده ابتدا یک برجستگی جوش مانند ( نودول ) که ناشی از تجمع سلول های پلاسمایی ، لنفوسیت و ماکروفاژهاست بوجود می آید .
شهرها اتفاق می افتد در محل جوش زخمی باندازه ۳-۱ سانتی متر شکل می گیرد که اطراف آن آماسی است انگل ها در زخم تکامل یافته پراکنده است مخصوصا در ناحیه حاشیه ای آن بهبودی در عرض چند ماه صورت می گیرد و ایمنی پایدار شکل می گیرد .
های لیشمانیا تروپیکا خشک و لیشمانیا ماژور مرطوب هستند هر دو تا ممکن است به عفونت های ثانویه آلوده گردند ( yaws ) از جمله یاوز ( یک بیماری پوستی غده غده ای شدن که ناشی از نوعی اسپیروکت است ) و میاز ( آلودگی زخم بالا رو حشرات ) بعد از بهبود زخم چه بصورت خود بخود و چه درمان یک ایمنی پایداری بوجود می آید. که ایجاد بیماری احشایی می کنند لیشمانیا گروه مکزیکی ( مکزیکان گروپ ) که ایجاد جراحت خود بهبودی یابنده پوستی می کند و گروه لیشمانیا برازیلنیسیس که ایجاد جراحات جلدی و مخاطی – جلدی می کند بیماری بصورت نقاشی و حک شده بر روی سفال های بدست آمده از پرو که متعلق به ۴۰۰ سال پیش از میلاد مسیح است نشاند داده شده است بیماری در تمام کشورهای آمریکایی لاتین بجز شیلی واروگوئه گسترش دارد و مواردی هم از تگزاس گزارش شده است بیماری یک بیماری مشترک بین انسان و حیوان است که میزبان مخزن در بین حیوانات وحشی دارد .
و اطراف آن التهابی است. لیشمانیا مکزیکانا ایجاد زخم شیکلروس می کند که می تواند تقریبا باعث تخریب تمام گوش خارجی شود.
این زخم ها ممکن است در عرض چندماه تا چند سال از آغاز عفونت بوجود آیند بیماری عمدتادرافرادیکه شاغل در جنگل هستند بروز می کند و موسوم به اسپاندیا می باشد تخریب جنگل ها موجب گرایش و تطابق بعضی از گونه های پشه خاکی در محیط های مسکونی و لذا دام اهلی بعنوان دومین میزبان های مخزن بشمار می آیند.
آشفتگی ( دفورمیتی ) اعضاء مشاهده می شود. اغلب بافت های استخوانی و غضروفی بینی و همچنین کام سخت و نرم از بین می روندومرگ بدنبال عفونت ثانویه یا سوء تغذیه رخ می دهد . و گاه بیماری به اشتباه جذام و یا دیگر عفونت های باکتریایی و قارچی تشخیص داده می شود.
باگرانولومای ماکروفاژی و ضخیم شدن پوست مشخص شده مشاهده میشود. این فرم مثل جذام لپرو ماتوزی و مثل بیماریهای قارچی است و گاه در تشخیص دچار اشتباه می کند. ● پراکندگی لیشمانیازیس کشور جز کشور های توسعه یافته جهان هستند تخمین زده می شود حدود ۳۵۰ میلیون نفر در معرض ابتلا به گونه های مختلف لیشمانیا هستند. بروز موارد جدید در سال ، حدود ۲ میلیون نفر است ( ۵/۱ میلیون لیشمانیای جلدی و نیم میلیون نفر لیشمانیای احشایی) مثل خیلی از بیماریهای گرمسیری عفونت های لیشمانیایی با توسعه اقتصادی ساخت بشر و تغییرات محیط اطراف توسط انسانها باعث افزایش تعارض ( در معرض قرار گرفتن ) انسان با پشه خاکی ناقل بیماری باشد برداشت الوار از جنگل ها استخراج معادن ، سدسازی ها ، تعریض اراضی برای کشت ، برنامه های جدید آبیاری ، جاده سازی و جنگل ها دست نخورده از جمله در آمازون ، گسترش مهاجرت های اقتصادی و شهرنشینی های فزاینده و سریع در سطح جهان علل اصلی افزایش موارد ابتلا به این بیماری است .
بعنوان اصلی ترین مخزن لیشمانیا تروپیکا در مرکز آسیاست و در بعضی از مناطق سگ وگربه وسنجاب ها نیزعفونی میگردند . در آمریکای لاتین جوندگان بعنوان مخزن برای گونه هایی از لیشمانیا هستند . اخیرا یک افزایش درلیشمانیای احشائی پدیدآمده که ناشی ازاشاعه جهانی ایدز میباشدعفونت توامان ایدز لیشمانیا رامیتوان یک بیماری وابسته یاگرایشی واقعی بحساب آوردمخصوصادراروپای جنوبی که۷۰-۲۵ درصدازلیشمانیای احشائی بالغین وابسته به عفونتHIV بوده است۹-۵/۱درصداز مواردایدزاز بیماری لیشمانیای احشائی تازه گرفتار شده ویادوباره فعال شده رنج می برند همه گیری هائی از لیشمانیا احشائی هنوز هم درسودان شرقی و جنوبی ودر تقاطع اریتره اتیوپی وسودان بروز می کند اخیرا بروز مجددی از لیشمانیای احشائی در برزیل رخ داده است.
سایت Oppedaes است.
بیماری ناشناخته ای شدند نشانه هایی که در آنها مشهود بود عبارت بودند از ضعف , خونریزی , از دست دادن موها و نارسایی کلیه , بیش از ۲۴ تا از سگ ها مردند . آزمایش مایعات مفصلی سگ های بیمار منجر به شناسایی انگل های لیشمانیایی شد و پس از آن تمام سگ های شکاری ( Fox hound ) آمریکا تحت آزمایش قرار گرفتند و ۱۲ درصد از آنها پادتن بر علیه لیشمانیا داشتند ولی نه در سگهای ولگرد میل بروک و نه سگهای ولگرد سایر نقاط آمریکا پادتنی مشاهده نشد .انتقال این بیماری مبهم بود .چون از چهارگونه پشه خاکی موجود در ایالات متحده هیچکدام به جز در نیوجرسی قادر به زندگی در شمال نیستند . دانشمندی در والتر راید چنین نظریه داد که سگها با تماسهای جنسی یا تماس زخمها ممکن است همدیگر را آلوده کرده باشند البته متخصصین لیشمانیا با این نظر موافق نیستند .
گزیده شده اند یک احتمال دیگر اینکه گونه ناشناخته ای از پشه خاکی در محل باشد یا حتی ناقلین دیگری وجود داشته باشد گونه لیشمانیا , لیشمانیا ایفانتوم است که به احتمال پایین بودن سطح اپیدمیکی آن در افراد ایالات متحده وجودش فراموش شده است.
انگلی و یا روش میکروسکوپی ا ست اخیراً PCR ژن های انگلی جهت تشخیص مورد استفاده قرار می گیرد.
با الگوهای معلوم در مقابل ژن های انگل روبرو میکنند . درمان بیماریهای لیشمانیایی هم بصورت موضعی و هم بصورت عمومی با استفاده از ترکیبات پنج ظرفیتی آنتیموان و ترکیبات معطره دیامیدین انجام میگیرد رایج ترین ترکیبات مورد استفاده از آنتیموان پنج ظرفیتی استیبو گلوکنات سدیم ( پنتوستام ) در آنتی مونیات ملگوسین (گلوکانتیم) که برای اولین با در سال ۱۹۴۷ و ۱۹۵۰ بکار گرفته شده است عدم توفیق در درمان و عود بیماری در تمام فرمهای لیشمانیازیس حدود ۲۵-۱۰ درصد موارد میباشد اگر این داروها موثر نبودند از پنتامیدین (لومیدین) و آمفوتریسین B)فانژیزون) که در سالهای ۱۹۴۰ و ۱۹۵۹ معرفی شده اند استفاده می شود .درمان لیشمانیای مخاطی – جلدی موفقیت آمیز نیست. گیرند لذا واکنش ایمنی هومورال بدلیل مخفی بودن انگل لیشمانیا شکل نمی گیرد از آنجائیکه انگل های درون سلولی در معرض واکنش هومورال میزبانی نیستند پادتن های گردش خون تاثیری بر عفونت ندارند وحتی ممکن است مضر نیز واقع شوند ایمنی بطور کلی وابسته به ایمنی سلول است و این براساس مشاهدات زیر است. با استفاده از سویه های همخون موش هایی که از نظر ژنتیکی مشخص و معلوم هستند آنهایی که در مقابل لیشمانیای ماژور جلدی بهبودی یافتند و آنهایی که بهبودی نیافتند موجب افزایش دانش ما در مورد واکنش ایمنی در برابر لیشمانیا است . میشود .
با تولید عامل نکروز توموری آنها (TNFalpha) که باعث تحریک در تولید آنزیم اکسید نیتریک سنستتاز می شود . نمود استعداد ابتلا به عفونت در موش های همخون حداقل بوسیله پنج جایگاه ژن کنترل می شود .
کمپلکس اصلی بافت رقابتی و NraMp۱ یا L Sh مشابه همین برای ژنهای مستعد کننده انسان نیز شناسایی شده است این ژن ها همچنین کنترل مقاومت دیگر پاتوژنهای درون سلولی مثل توکسوپلاسما , سل و جذام را بعهده دارند.
هومورال جلوگیری میکند. درحالیکه سلولهای Th۲ باعث فعال شدن ایمنی هومورال و از ایمنی وابسته سلولی جلوگیری میکند.
چند منطقه آزمایش شده است . در برزیل ( لیشمانیا امازونیسیس ) و در ونزوئلا ( لیشمانیا مکزیکن و لیشمانیا برازیلنسیس ) به ارتش اشغالگر اسرائیل نیز توصیه شده است که سربازانشان برعلیه لیشمانیا واکسینه کنند علاوه بر این بعضی از واکسنهای مولکولی ( واکسن های افتراقی ) که برای آزمایش انتخاب شده اند : GP۶۳ که مهمترین گلیکوپروتئین سطحی لیشمانیا و LPG ( لیپو فسفو گلیکان ) سطحی پارازیت است. واکسن های DNA که تجلی gp۶۳ را دارد معلومشده است تا حدودی موثر است.
میزبانان پستاندار افزایش پیدا میکند , آنها توانستند این فرایند را در کشت انگل تقلید کند. بطوریکه مرحله Log ) فاز لگاریتم ) غیرعفونی و مرحله خاموشی یا تثبیت عفونی بوده است. آنها این مراحل را به ترتیب پیش چرخه ای (Procyolic ) و فوق چرخه ای (Metacyolic ) پروماستیگوت نامیدند. پروماستیگوت های متاسیکلی لیشمانیا ماژور را می توان با عدم توانایی آگلوتینه کردن لکتین , آگلوتیمین بادام زمینی تشخیص داد .
لیشمانیای ماژور عفونی و غیرعفونی .دو گالاکتوز متصل به لکتین و آگلوتینین بادام زمینی ( PNA ) و آگلوتنین کرچک (RCA ) می توانند ۱۰۰% پروماتیسگوت های فاز Log را آگلوتینه نماینددر حالیکه فقط ۵۰ در صد پروماستیگوت هامرحله تثبیت راآگلوتینه می کنند افزودن mMD ۱۰گالاکتوز مانع از آگلوکتیناسیون میشود.
|
|
حمیده رحیمی Visceral Leishmaniasis (Kala-Azar)Clinical ManifestationsLike cutaneous leishmaniasis, visceral leishmaniasis begins with a nodule at the site of inoculation. This lesion rarely ulcerates and usually disappears spontaneously in a few weeks or months. In contrast to cutaneous leishmaniasis, symptoms and signs of systemic disease develop, such as undulating fever, malaise, diarrhea, splenomegaly, hepatomegaly, lymphadenopathy, emaciation, anemia, and leukopenia (Fig. 82-12). Infiltrative or nodular lesions of the skin may appear after treatment (post-kala-azar dermal leishmaniasis), a condition seen frequently in India. In some areas of Europe and Latin America, L infantum may cause a cutaneous form without apparent visceral involvement (Fig. 82-13). Subclinical cases also occur. FIGURE 82-12 Visceral leishmaniasis in a child from Honduras with marked emaciation and hepatosplenomegaly. (Courtesy of Carlos Ponce.) FIGURE 82-13 Nodular cutaneous leishmaniasis produced by L infantum in Costa Rica. Structure, Multiplication and Life CycleThe Leishmania species that cause kala-azar are similar in morphology and life cycle to other members of the same genus. Classification and Antigenic TypesKala-azar can be caused by at least three Leishmania species, which are differentiated by zymodeme, serodeme, and DNA hybridization. Leishmania donovani and L infantum are responsible for visceral leishmaniasis in the Old World; L chagasi and L infantum (which may be the same organism) cause the disease in the New World. PathogenesisIn more serious cases of visceral leishmaniasis the parasites, which can resist the internal body temperature, invade internal organs (liver, spleen, bone marrow, and lymph nodes) where they occupy the reticuloendothelial cells. The pathogenetic mechanisms of the disease are not fully understood, but, clearly, in those organs that exhibit marked cellular alteration, hyperplasia of histiocytes leads to hypertrophy. Parasitized macrophages replace hematopoietic tissue in the bone marrow. Patients with advanced disease are prone to superinfection with other organisms.
Host DefensesHigh levels of IgG and other immunoglobulins are common. The Montenegro skin test for delayed hypersensitivity is usually negative during the disease but becomes positive after treatment. EpidemiologyIn India, transmission occurs in villages in an anthroponotic man-sand fly-man cycle without nonhuman reservoir. In Europe and Africa, several rodents may act as reservoirs. In rural semiarid zone of Latin America, both wild and domestic dogs enter the epidemiological chain and the vector is a common anthropophilic and zoophilic sand fly, Lutzomyia longipalpis, abundant in and around houses. The disease is more common in children in both Latin America and the Mediterranean area. DiagnosisThe typical symptoms, particularly hepatosplenomegaly and the pancytopenia, strongly suggest visceral leishmaniasis. The parasite usually can be demonstrated in stained or cultured bone marrow or spleen material. Serologic tests (ELISA, immunofluorescent antibody) are useful, particularly in surveys. ControlThe same insecticides and drugs that work for cutaneous leishmaniasis are used for visceral leishmaniasis. Aromatic diamidines are also used. |



Toxoplasma Gondii
J. P. Dubey
General Concepts
Clinical Manifestations
Infection is often asymptomatic. Immunocompetent individuals may present with fever, lymphadenopathy, muscle aches, and headache. Congenitally infected children may suffer impaired vision and mental retardation. Immunosuppressed patients may have central nervous system disease (encephalitis).
Structure and Life Cycle
Members of the cat family (Felidae) are the definitive hosts; many mammals and birds serve as intermediate hosts. Infection is contracted by ingesting either oocysts or meat containing live organisms. Organisms enter the intestinal epithelium and can spread to many host tissues. Individual organisms are lunate, about 6 X 2 µm, and multiply within host cells. Tissue cysts containing hundreds of quiescent organisms may form as infection wanes. Toxoplasma reproduces sexually only in cats. Organisms infecting the intestinal epithelium produce oocysts which are shed in the feces. Mature oocysts are approximately 12 µm in diameter and contain eight infective sporozoites.
Classification and Antigenic Types
Toxoplasma gondii, a member of the Apicomplexa, is the sole species.
Multiplication and Life Cycle
A sexual multiplication by cell division can occur in virtually any host cell.
Pathogenesis
Host cells are destroyed by active multiplication of T gondii. Necrotic foci may result. Congenital infection often involves the retina and brain; focal chorioretinitis may result in impaired vision. Brain involvement in immunosuppressed patients may lead to large necrotic abscesses. Disease reactivation in immunosuppressed patients may result from the rupture of a tissue cyst.
Host Defenses
Immunocompetent individuals mount an effective cell mediated immune response that eradicates active infection within weeks or months and results in immunity against reinfection. Tissue cysts are unreactive and may persist for the life of the host.
Epidemiology
Toxoplasmosis shows a nonseasonal worldwide distribution. Most natural infections are acquired by ingesting undercooked meat containing tissue cysts or food contaminated by cat feces.
Diagnosis
Diagnosis is based on serology and on histologic examination of tissues.
Control
Infection may be prevented by thorough cooking of meat and by proper management of cats. Acute cases are treated with sulfadiazine and pyrimethamine.
INTRODUCTION
Toxoplasma gondii is an intestinal coccidium that parasitizes members of the cat family as definitive hosts and has a wide range of intermediate hosts. Infection is common in many warm-blooded animals, including humans. In most cases infection is asymptomatic, but devastating disease can occur.
Clinical Manifestations
Toxoplasma gondii usually parasitizes both definitive and intermediate hosts without producing clinical signs. In humans, severe disease is usually observed only in congenitally infected children and in immunosuppressed individuals, including patients with acquired immune deficiency syndrome (AIDS). Postnatally acquired infections may be local or generalized and are rarely severe in immunocompetent individuals. Lymphadenitis is the most common manifestation in humans. Any node can be infected, but the deep cervical nodes are the most commonly involved. Infected nodes are tender and discrete but not painful; the infection resolves spontaneously in weeks or months. Lymphadenopathy may be accompanied by fever, malaise, fatigue, muscle pains, sore throat, and headache.
Encephalitis is an important and severe manifestation of toxoplasmosis in immunosuppressed patients including patients with AIDS. Symptoms may include headache, disorientation, drowsiness, hemiparesis, reflex changes, and convulsions. Coma and death may ensue.
Prenatally acquired T gondii often infects the brain and retina and can cause a wide spectrum of clinical disease. Mild disease may consist of slightly diminished vision, whereas severely diseased children may exhibit a classic tetrad of signs: retinochoroiditis, hydrocephalus, convulsions, and intracerebral calcifications. Hydrocephalus is the least common but most dramatic lesion of congenital toxoplasmosis (Fig. 1). Ocular disease is the most common sequela.

FIGURE 84-1 Girl with hydrocephalus due to congenital toxoplasmosis. (From Dubey JP, and Beattie CP. Toxoplasmosis of animals and Man. CRC Press, Baca Raton, Florida, 52, 1988.)
Toxoplasma gondii is capable of causing severe disease in animals other than humans. It is one of the major causes of abortion in sheep and goats in many countries, including Australia and the United States. It is important to diagnose toxoplasmic abortion to distinguish it from other causes of abortion, because congenital transmission of T gondii occurs only during the initial infection of the mother and the animal is safe for breeding thereafter. Cats, dogs, and many other pets can die of pneumonia, hepatitis, and encephalitis due to toxoplasmosis. In dogs, clinical toxoplasmosis is often associated with concurrent distemper virus infection. Certain species of marsupials and New World monkeys are highly susceptible to toxoplasmosis.
Structure, Multiplication, and Life Cycle
The life cycle of T gondii was described only in 1970, when it was discovered that the definitive hosts are members of the family Felidae, including domestic cats. Various warm-blooded animals serve as intermediate hosts. Toxoplasma gondii is transmitted by three known modes: congenitally, through the consumption of uncooked infected meat, and via fecal matter. Figure 2 shows the life cycle of T gondii.

Figure 84-2 Life cycle of Toxoplasma gondii. Cats, the definitive hosts of T gondii, can become infected by ingesting sporulated oocysts or (most often) infected animals. The oocysts are infectious to most mammals and birds. Toxoplasma can be transmitted to intermediate hosts through oocysts, by carnivorism, or transplacentally. Transplacental transmission is most important in humans and sheep. (From Dubey JP, Toxoplasmosis. J Am Vet Med Assoc 189:166, 1986.)
Cats acquire Toxoplasma by ingesting any of three infectious stages of the organism: the rapidly multiplying forms called tachyzoites (Fig. 3), the quiescent bradyzoites that occupy cysts in infected tissue (Fig. 4), and the oocysts shed in feces (Fig. 5). Successful infection of the cat is revealed by the shedding of oocysts in the feces. The chance of infection and the prepatent period (the time between infection and the shedding of oocysts) varies with the stage of T gondii ingested. Fewer than 50 percent of cats shed oocysts after ingesting tachyzoites or oocysts, whereas nearly all cats shed oocysts after ingesting tissue cysts. Only the cyst-induced cycle has been studied in detail.

FIGURE 84-3 Tachyzoites of T gondii.
A. Extracellular (arrow) released from host cells. Compare their size with red blood cells and a lymphocyte. Impression smear, Giemsa stain. Bar = 20 µm.
B. Intracellular in cell culture. Note a group arranged in a rosette (arrow) and vacuole (arrowhead) around a tachyzoite. Immunohistochemical stain with a tachyzoite-specific monoclonal antibody. Bar = 20 µm.
C. Transmission electron micrograph of an intracellular tachyzoite. Note a parasitophorous vacuole (PV) around the tachyzoite. Parasite organelles visible in this picture include a conoid (c), micronemes (m), dense granules (dg) nucleus (n) and rhoptries (r). Bar=0.8 µm. (Courtesy of Dr. D.S. Lindsay, Auburn University, AL.)

FIGURE 84-4 Tissue cysts of T gondii.
A. Tissue cyst freed from mouse brain. Note a thin (arrow) cyst wall enclosing hundreds of bradyzoites. Unstrained. Bar = 20 µm.
B. Two tissue cysts (arrows) in section of brain. Hematoxylin and eosin stain. Bar = 20 µm.
C. Transmission electron micrograph of a small tissue cyst in cell culture. Note thin cyst wall (arrow) enclosing 6 bradyzoites (arrowheads). Bar = 1.0 µm. (Courtesy of Dr. D.S. Lindsay, Auburn University, Auburn, AL.)
When a cat ingests meat containing tissue cysts, the cyst wall is dissolved by the proteolytic enzymes in the stomach and small intestine, releasing the bradyzoites. The bradyzoites, which are a slow multiplying stage, penetrate the epithelial cells of the small intestine and initiate the formation of numerous asexual generations before the sexual cycle (gametogony, the production of gametes) begins (Fig. 5A). After the male gamete (Fig. 5B) fertilizes the female gamete, two walls are laid down around the fertilized zygote to form the oocyst, which is excreted in the feces in an unsporulated stage (Fig. 5C).

FIGURE 84-5 Sexual states of T gondii
A. Schizonts (double arrowheads), female gamonts (arrows), and male gamonts (arrowheads) in section of superficial epithelial cells of the small intestine of a cats. Hematoxylin and eosin stain. Bar = 15 µm.
B. Three male gametes each with 2 flagella (arrowheads) compared with a merozoite (arrow). Impression of intestinal epithelium of a cat. Giemsa stain. Bar = 10 µm.
C. Unsporulated oocytes (arrowheads) in feces of a cat. Note 2 oocysts of another feline coccidium, Isospora felis (arrowheads). Isospora felis sporulates faster than T gondii. The oocysts on top of the picture already contains 2 sporocysts while all T gondii oocysts are unsporulated. Unstained. Bar - 65 µm.
D. Transmission electron micrograph of a sporulated oocyst. Note thin oocyst wall (arrow), 2 sporocysts (arrowheads) and 4 sporozoites (double arrowheads) in sporocysts. Bar - 2.25 µm. (Courtesy of Dr. D.S. Lindsay, Auburn University, Auburn, AL.)
Oocysts measure approximately 10 by 12 µm. Sporulation occurs outside the body, and the oocyst becomes infectious 1 to 5 days after excretion. Each sporulated oocyst contains two sporocysts and each sporocyst contains four sporozoites (Fig. 5D). Sporulated oocysts are remarkably resistant and can survive in soil for several months.
At the same time that some bradyzoites enter the surface epithelial cells of the feline intestine and multiply there to produce oocysts, other bradyzoites penetrate the lamina propria and begin to multiply as tachyzoites. Tachyzoites are about 6 X 2 µm in size and generally lunate (Fig. 3A). Within a few hours of infection, tachyzoites may disseminate to extraintestinal tissues through the lymph and blood. Tachyzoites can enter almost any type of host cell (Fig. 3C) and multiply until the host cell is filled with parasites and dies (Fig. 3B). The released tachyzoites enter new host cells and multiply. This cycle may result in microfoci of tissue necrosis. The host usually overcomes this phase of infection, and the parasite then enters the "resting" stage in which bradyzoites are isolated in tissue cysts. Tissue cysts are formed most commonly in the brain, liver, and muscles. Cysts in neural tissues are up to 60 µm in diameter and contain hundreds of bradyzoites in a thin membrane (Fig. 4). Tissue cysts usually cause no host reaction and may remain for the life of the host.
In nonfeline intermediate hosts, such as humans or mice, the extraintestinal cycle of T gondii is similar to the cycle in cats. However, sexual stages are produced only in the feline definitive hosts.
Pathogenesis
Most cases of toxoplasmosis in humans are probably acquired by the ingestion of either tissue cysts in infected meat or oocysts in food contaminated with cat feces. Bradyzoites from the tissue cysts or sporozoites released from oocysts penetrate the intestinal epithelial cells and multiply in the intestine. Toxoplasma gondii may spread both locally to mesenteric lymph nodes and to distant organs by invading the lymphatics and blood. Necrosis in intestinal and mesenteric lymph nodes may occur before other organs become severely damaged. Focal areas of necrosis may develop in many organs. The clinical picture is determined by the extent of injury to these organs, especially to vital and vulnerable organs such as the eye, heart, and adrenals. Toxoplasma gondii does not produce a toxin; necrosis is caused by intracellular multiplication of tachyzoites.
Opportunistic toxoplasmosis in AIDS patients usually represents reactivation of chronic infection. The predominant lesion of toxoplasmosis - encephalitis in these patients is necrosis, which often results in multiple abscesses, some as large as a tennis ball (Fig. 6).

FIGURE 84-6 Section of brain from an AIDS patient with fatal toxoplasmosis. Note a large focus of necrosis, 2 tissue cysts (arrows) and numerous tachyzoites (arrowheads - all black dots are tachyzoites). Immunohistochemical stain with anti-T gondii serum Bar - 100 µm.
Host Defenses
The host may die from toxoplasmosis but much more often recovers and acquires immunity. Inflammation usually follows necrosis. By about the third week after infection, Toxoplasma gondii tachyzoites begin to disappear from visceral tissues and may localize as tissue cysts in neural and muscular tissues. Toxoplasma tachyzoites may persist longer in the spinal cord and brain because immune responses are less effective in these organs. Chronic infections may be reactivated locally (for example, in the eye). Reactivation possibly results from the rupture of a tissue cyst. Probably tissue cysts rupture periodically during the life of the host, and the bradyzoites released are normally destroyed by the host's immune responses. This reaction may cause local necrosis accompanied by inflammation. Hypersensitivity is said to play a major role in such reactions; however, in immunocompetent hosts the infection usually subsides, with no local renewed multiplication of Toxoplasma. In immunosuppressed patients, rupture of a tissue cyst may result in renewed multiplication of bradyzoites into tachyzoites, and the host may die from toxoplasmosis. The cause of cyst rupture is not known. Chronic latent T gondii infection can be experimentally reactivated by excessive doses of corticosteroids, antilymphocyte serum and other immunosuppressive therapies.
Epidemiology
Toxoplasma gondii infection in humans is widespread throughout the world. Approximately half a billion humans have antibodies to T gondii. The incidence of infection in humans and animals may vary in different parts of a country. The cause for these variations is not yet known: environmental conditions, cultural habits, and animal species are among factors that may determine the degree of natural spread of Toxoplasma gondii. Only a small proportion (less than 0.1 percent) of people acquire infection congenitally. Immunocompetent mothers of congenitally infected children do not give birth to infected children in subsequent pregnancies. However, repeated congenital infection can occur in mice, rats, guinea pigs, and hamsters without reinfection from outside sources.
The relative frequency with which postnatal toxoplasmosis is acquired by eating raw meat and by ingesting food contaminated by oocysts from cat feces is unknown and difficult to investigate. Both modes of infection are reported to cause clinical toxoplasmosis. Toxoplasma gondii infection occurs commonly in many animals used for food (for example, sheep, goats, pigs, and rabbits). Infection is less prevalent in cattle than in sheep or pigs.
Oocysts are shed by cats - not only the domestic cat - but also other felids such as ocelots, margays, jaguarundi, bobcats, Pallas cats, and Bengal tigers. However, oocyst formation is greatest in the domestic cat. Widespread natural infection is possible because a cat may excrete millions of oocysts after ingesting few tissue cysts. Oocysts are resistant to most ordinary environmental conditions and can survive in moist conditions for months and even years. Invertebrates such as flies, cockroaches, and earthworms can spread oocysts mechanically.
Only a few cats may be involved at any one time in spreading T gondii in an area: at any given time as little as 1 percent of the domestic cat population in the United States is shedding oocysts. It is not known whether cats shed oocysts only once or several times in nature. Under experimental conditions, cats usually do not shed oocysts after reinoculation with T gondii tissue cysts, but this immunity to T gondii in cats does wane with time.
Diagnosis
Diagnosis of toxoplasmosis can be aided by serologic or histocytologic examination. Clinical signs of toxoplasmosis are nonspecific and cannot be depended on for a definite diagnosis; toxoplasmosis clinically mimics several other infectious diseases.
Many serologic tests have been used to detect antibodies to T gondii. The most reliable of these is the Sabin-Feldman dye test. Live virulent tachyzoites of T gondii are used as antigen and are exposed to dilutions of the test serum and to a complement accessory factor resembling complement that is obtained from Toxoplasma-antibody free-human serum. This test is sensitive and so far is the most specific test for toxoplasmosis. Its main disadvantages are its high cost and the human hazard of using live organisms.
The indirect fluorescent antibody test (IFAT) overcomes some of the disadvantages of the dye test. In IFAT, killed tachyzoites of Toxoplasma, which are available commercially, are used as antigen. Titers obtained by IFAT are similar to those from the dye test. Disadvantages of the IFAT are that a microscope with UV light is needed, fluorescent anti-species globulin is required for each species to be tested, and false-positive titers may occur in hosts with anti-nuclear antibodies. The suitability of IFAT in animal diagnostic work is therefore limited, but it has proved useful in diagnosing acquired human toxoplasmosis. Other serologic tests including the indirect hemagglutination test, the latex agglutination test, modified agglutination test, and the enzyme-linked immunoabsorbent assay (ELISA), offer some advantages. For example, agglutination tests are easy to perform.
Soluble antigens used for indirect hemagglutination tests are now commercially available in several countries, including the United States. Although this test is easy to perform, it usually does not detect antibodies during the acute phase of toxoplasmosis. In the modified agglutination test, whole killed tachyzoites are used as antigen, and the test serum is treated with 2-mercaptoethanol to eliminate nonspecific agglutinins. The ELISA test using soluble antigens appears to be specific and may become the standard test in the future.
A single positive serum sample proves only that the host has been infected at some time in the past. Serologic evidence for an acute acquired infection is obtained when antibody titers rise by a factor of 4 to 16 in serum taken 2 to 4 weeks after the initial serum collection, or when specific IgM antibody is detected. The finding of antibody in even undiluted serum is useful in the diagnosis of ocular toxoplasmosis because patients with this disorder usually have low T gondii antibody titers.
Diagnosis can be made by finding T gondii in host tissue removed by biopsy or at necropsy. This procedure is particularly useful in immunosuppressed patients or patients with AIDS, in whom antibody synthesis may be delayed and low. Toxoplasma gondii infection can be rapidly diagnosed by making impression smears of lesions on glass slides. After drying for 10 to 30 minutes, the smears are fixed in methyl alcohol and stained with Giemsa stain. Well-preserved T gondii organisms are crescent-shaped and stain well with any of the Romanowsky stains (Fig. 3A); however, degenerating organisms common in lesions usually appear oval and have cytoplasm that stains poorly compared to the nucleus. A diagnosis of toxoplasmosis should not be made unless organisms with the typical structure are seen, as degenerating host cells may resemble degenerating T gondii parasites. In thin sections the tachyzoites are oval to round and usually do not stain differently from host cells. Tissue cysts are occasionally encountered in areas with lesions (Fig. 4B). Tissue cysts are usually spherical and have silver-positive walls; the bradyzoites stain strongly with periodic acidSchiff stain. Immunohistochemical staining and polymerase chain reaction (PCR) can be used to identify T gondii tissue cysts or tachyzoites in tissues, even those fixed in formalin. Electron-micrographic examination can aid in diagnosis (Fig. 3C). Computed tomography techniques are also useful in the diagnosis of human cerebral toxoplasmosis. Inoculation of biopsy materials into laboratory mice and/or cell cultures can help diagnosis.
Control
Sulfonamides and pyrimethamine (Daraprim) are two drugs widely used to treat toxoplasmosis in humans. They act synergistically by blocking the metabolic pathway involving p-aminobenzoic acid and the folic-folinic acid cycle, respectively. These two drugs usually are well tolerated by the patient, but sometimes thrombocytopenia, leukopenia, or both may develop. These effects can be overcome without interrupting treatment by administering folinic acid and yeast because the vertebrate host can utilize presynthesized folinic acid, whereas T gondii cannot. The commonly used sulfonamides, sulfadiazine, sulfamethazine, and sulfamerazine, are all effective against toxoplasmosis. Generally, any sulfonamide that diffuses across the host cell membrane is useful in antitoxoplasmid therapy. Although these drugs are helpful when given in the acute stage of the disease, usually they will not eradicate infection when active multiplication of the parasite occurs. Because sulfa compounds are excreted within a few hours of administration, they must be administered in daily divided doses. Spiramycin, a drug used in France to treat pregnant women to minimize the effects of congenital toxoplasmosis, is not approved for toxoplasmosis in the United States. As yet, there are no effective drugs to kill tissue cysts.
No killed vaccine is currently available to reduce or prevent congenital infections in humans and animals, but research to develop such an agent is under way. A live vaccine using a nonpersistent T gondii strain is available in Europe and New Zealand to reduce abortion in sheep.
To prevent T gondii infection, several precautions should be taken. Meat should be cooked to 66°C throughout before eating. Hands should be washed with soap and water after handling meat. Raw meat should never be fed to cats; only dry or canned food or cooked meat should be fed. Cats should be kept indoors and litter boxes changed daily. Cat feces should be flushed down the toilet or burned. Litter pans should be cleaned by immersing them in boiling water. Gloves should be used while working in the garden. Children's sandboxes should be covered when not in use.
REFERENCES
Dubey JP, Beattie CP: Toxoplasmosis of Animals and Man. CRC Press, Boca Raton, FL, 1988
Gazzinelli RT, Denkers EY, Sher A.: Host resistance to Toxoplasma gondii: model for studying the selective induction of cell-mediated immunity by intracellular parasites. Infect Agent Dis 2:139, 1993
Georgie VA: Management of toxoplasmosis. Drugs, 48:179, 1994
Guerina NG, Hsu HW, Meissner HC, Meissner HC, et al. Neonatal serologic screening and early treatment for congenital Toxoplasma gondii infection. N Eng J Med, 330:1858, 1994
Remington, JS, McLeond R, Desmonts G.: Toxoplasmosis. In: Remington JS, Klein JO, (eds.): Infectious Diseases of the Fetus and Newborn Infant. Philadelphia: W.B. Saunders Company, 1995
Injections
Various routes exist for injecting mice. [Click the image below to see a chart] 
It is important that you discuss the appropriate route, volume, site and needle selection with your veterinarian.
All injections must be described in your approved Animal Study Proposal.
All injections must be performed using sterile needles and syringes.
A new needle and syringe should be used for each cage of mice.
 Intramuscular Injections
Intramuscular Injections
Regardless of the method used for intramuscular injections, it must be noted that the sciatic nerve runs along the length of the femur.
It is very important to avoid injuring this nerve.
This is best accomplished by pointing the needle, caudally rather than cranially, into the caudal thigh muscles.
 It is imperative that the mouse be properly restrained.
It is imperative that the mouse be properly restrained.
If the mouse is allowed to kick or struggle, this could cause injury to the muscles or the nerve.

It is best to swab the area with 70% ethanol before placing the needle and to aspirate to look for blood before injecting.
One of the following methods may be used:

Method #1
Restrain the mouse by the scruff method.
Secure the rear foot nearest to you beneath your little finger and lower thumb.
Swab the area to be injected with 70% ethanol.

Insert the needle, bevel up, into the caudal thigh at a 45° angle.
Aspirate to ensure that you have not entered a blood vessel.
If no blood is seen, slowly inject the material.
 Method #2
Method #2
Restrain the mouse by the scruff method.
Swab the area to be injected with 70% ethanol.
Insert the needle into the caudal thigh, bevel up.
Aspirate and inject.
Since the foot has not been secured, make sure the mouse does not kick.
 Method #3
Method #3
A technique using two people can also be used for IM injections.
One person restrains the mouse by the scruff method with one hand and steadies the leg to be injected with the other.
The second person identifies the caudal thigh muscles, swabs the area with 70% ethanol, aspirates and injects the material.
 Method #4
Method #4
A tail first restrainer can also be used for IM injections.
Gently pull the foot of the leg to be injected through the restrainer and locate the caudal thigh muscle.
Hold the foot firmly, swab the site with 70% ethanol, aspirate and inject the material with the needle bevel up.
 Subcutaneous Injections
Subcutaneous Injections
Restrain the mouse by the scruff method. Use your thumb and forefinger to make a tent of skin over the scruff.
Prep the area with 70% ethanol.
Insert the needle, bevel up, at the base of the tent.
The needle should be inserted parallel to the skin and should be directed toward the posterior of the animal.
Aspirate to ensure proper placement and inject the material.
Revised 4/6/04 9
 Intraperitoneal Injections
Intraperitoneal Injections
Restrain the mouse by the scruff method. Expose the ventral side of the animal, tilting the head down at a slight angle.
Prep the site with 70% ethanol.
The sterile needle should be placed, bevel up, in the lower right or left quadrant of the animal’s abdomen.
Insert the needle at a 30° angle.
Aspirate to ensure proper placement and inject the material.
 Intravenous Injections
Intravenous Injections
Warm the mouse under a heat lamp or other heating device, being sure not to OVERHEAT the animal.
The temperature should not exceed 85 - 90° Fahrenheit at the level of the animal.
Remove the mouse from the heat source immediately should any change in respiration rate or excessive salivation be observed.
Other heating devices, such as disposable handwarmers, may be used in lieu of a heat lamp.
 Place the animal in a restraint device and stabilize the tail between the thumb and forefinger of the hand that will not be manipulating the syringe.
Place the animal in a restraint device and stabilize the tail between the thumb and forefinger of the hand that will not be manipulating the syringe.
Prep the tail with 70% ethanol.
Attempt the injection starting at the middle or slightly distal part of the tail.
With the tail under tension, insert the needle, bevel up, approximately parallel to the vein and insert the needle at least 3 mm into the vein.
DO NOT ASPIRATE, as it will cause the vein to collapse.
Inject the material in a slow, fluid motion.
You should be able to see the vein blanch if the needle is properly positioned.
If any swelling at the injection site or resistance to injection occurs, remove the needle and reinsert it slightly above the initial injection site.
 Intradermal Injections
Intradermal Injections
In order to perform intradermal injections, the mouse should be anesthetized.
Shave or pluck an injection site on the back of the animal to remove the hair.
Swab the site with 70% ethanol.
Insert the needle into the skin, bevel up, holding the needle nearly parallel to the plane of the skin.
Do not aspirate.
Inject the material.
The volume of the injection should be limited to 50 ىl per site to avoid tissue trauma.
 A properly performed intradermal injection will result in a small, round skin welt.
A properly performed intradermal injection will result in a small, round skin welt.
 Oral Gavage
Oral Gavage
Gavaging is used to dose an animal with a specified volume of material directly into its stomach.
Only a specialized, commercially available gavage needle should be used for attempting this procedure.
Fill the syringe with the appropriate volume of material and attach the needle.
Restrain the animal by the scruff.
Place the tip or ball of the needle into the animal’s mouth.
Slide the tip gently past the back of the tongue.
The needle should slide easily down the esophagus, if properly placed. DO NOT FORCE!!!
If any resistance is met, remove the needle and reinsert.
Do not aspirate.
Once the needle is properly placed, administer the material.
A table of injection routes, sites, volumes and needle sizes can be reached through the link below.
INJECTION SITES AND VOLUMES [Click this link to see a chart]
Revised 4/6/04 10
-
Select the appropriate restraint method, injection route, volume, site and needle gauge for your research purposes.
-
Prep the site with 70% ethanol.
-
Always inject with the needle bevel up.
-
When warming animals, DO NOT OVERHEAT.
-
All injection procedures must be described in your Animal Study Proposal.
-
Consult your veterinarian for further information concerning injections.
Remember
Source: US National Institutes of Health
Intravenous Injection in the Mouse
Intravenous administration of material can be technically difficult, and use of a restraining device is often required. These should be selected carefully to be an appropriate size for the animal to be injected. Too small a device can result in injuries to the animal, and can interfere with respiratory movements. Too large a restrainer can also result in injury, caused by movements during restraint.

After use, restraining devices should be cleaned thoroughly, to avoid pheromonally-induced stress or cross-infection.
Although rapid buffering by the blood and rapid dilution can allow a wider range of materials to be administered, injection into a peripheral vein can result in irritation, and in some circumstances flushing with normal saline should be considered.
It is common practice to stimulate dilation of the tail veins in mice either by placing them in a warmer environment (e.g. at 28-30 ºC) for up to 30 minutes, or by placing the tail in warm (30-35 ºC) water. If a warming box or incubator is used, its temperature should be monitored carefully. It is good practice to place an electronic thermometer adjacent to the animal’s cage in the incubator, as the temperature registered by the device thermostat may not be accurate.
Click to download the trainers pack for this technique which includes the video above (110Mb).
بررسی اثر اپیوم موضعی در درمان سالك
دكتر محمدعلی ماپار ، دكتر حسین كاوسی ، دكتر محمد علی دباغ
1-استادیار گروه پوست، 2- دستیار گروه پوست، 3- استادیار گروه داروسازی؛ دانشگاه علوم پزشكی اهواز
مقدمه
سالك بیماری شایع پوستی است كه در بسیاری از نقاط دنیا مشاهده میگردد. كشور ایران از جمله مناطقی است كه این بیماری به صورت بومی در آن وجود دارد(1). خوزستان از كانونهای مهم سالك در ایران است(2). عامل بیماری سالك در كشور ما معمولاً لیشمانیا تروپیكا و لیشمانیا ماژور و ناقلین آن فلبوتوموس پاپاتاسی، فلبوتوموس سرژانتی و فلبوتوموس انصاری میباشد(1).
سالك با نماهای بالینی مختلف تظاهر مینماید. ضایعة معمولاً به صورت پاپول قرمز رنگی شروع میگرددكه به تدریج بزرگتر میشود و در نهایت به صورت پلاك زخمی درمیآید و در طی حدود یك سال با برجای گذاشتن جوشگاه بدنما بهبود مییابد(3).
سالك یك بیماری خود محدود شونده است ومعمولاً درطی یك سال بهبود مییابد. آنچه كه درمان سالك را ضروری میسازد این است كه در 10% موارد، بیماری سالك به صورت مزمن یا لوپوئید درمیآید(3). وجود ضایعه درنواحی غیرپوشیده بدن منجر به جوشگاه بدنما میگردد. در كانونهای شهری، انسان ممكنست مخزن بیماری باشد و با درمان مناسب موارد انسانی، از احتمال انتقال عفونت به افراد سالم كاسته میشود(3). تاكنون روشهای مختلفی برای درمان بیماری سالك به كار رفته و در مطالعات متعدد، نتایج متفاوتی از اثرات این داروها گزارش گردیده است.
اپیوم خام یا تریاك معمولاً به عنوان ضد درد كاربرد دارد. این ماده دارای اثرات دیگری در درمان سرفه، اسهال ساده و در گذشته در بیماری دیابت بوده است(4). كپسول خشخاش به صورت موضعی جهت تسكین درد و التهاب ساده و همچنین به صورت بخور در لارنژیت مصرف گردیده است(4).
در مطالعهای بر روی موشهای آزمایشگاهی Balb/c كه به صورت تجربی با لیشمانیاماژور آلوده و زخم سالكی در آنها ایجاد گردیده بود، از اپیوم موضعی روزانه یك مرتبه به مدت دو هفته استفاده گردید كه در پایان دورة درمان 5/87% موشها از نظر بالینی بهبود یافته و 7/68% هم از نظر بالینی بهبودی نشان داده و هم ضایعات از نظر وجود جسم لیشمن در آزمایش مستقیم منفی شده بودند. روند بهبودی موشهای درمان شده به مدت 6 ماه پیگیری گردید و در این مدت هیچ گونه زخمی در ناحیة درمان یافته و سایر نقاط بدن حیوان مشاهده نگردید(5).
در بعضی مناطق روستایی ایران كه از نظر سالك اندمیك میباشد از تریاك بصورت موضعی در درمان سالك استفاده میگردد. این افراد ادعا نمودهاند كه در طی مدت كوتاهی بطور كامل بهبود یافتهاند. با توجه به استقبال بیماران از داروهای موضعی، هزینه پایین و اثربخشی دارو در مدلهای حیوانی، تلاش شد كه اثرات اپیوم موضعی در درمان سالك در انسان در یك مطالعه كارآزمایی بالینی
دوسوكور مورد بررسی قرار گیرد.
روش اجرا
این مطالعه به صورت دوسوكور در مركز پیشگیری و مبارزه با بیماریهای واگیر در شهرستان اهواز انجام گرفت. مدت مطالعه دو سال و در سالهای 79-1378 بود. بیمارانی كه در این مطالعه قرار گرفتند دارای معیارهایی شامل: سن بالای دو سال، تعداد ضایعات كمتراز 6 عدد، دورة بیماری كمتر از 3 ماه، عدم مصرف داروی مؤثر بر سالك قبل از مراجعه و مثبت بودن آزمایش مستقیم(اسمیر) از نظر وجود جسم لیشمن بودند. زنان مبتلا به سالك كه شیرده یا حامله بودند وارد مطالعه نشدند.
بیماران پس از دادن آگاهی مختصر در مورد سالك و نوع درمان مورد نظر و كسب موافقت بیماران یا والدین آنهاتحت مطالعه قرار میگرفتند. از 96 بیمار واجد شرایط كه در مطالعه قرار گرفتند 58 بیمار دورة درمان را به پایان رساندند و 38 بیمار به دلایل مختلف از مطالعه خارج شدند.
بیماران در دو گروه دریافت كننده دارو یا دارونما بر اساس ظرف دارویی كه از قبل كدگذاری گردیده بود تقسیم گردیدند. در روزهای زوج ظرف دارویی با كدA و در روزهای فرد ظرف دارویی با كد B به بیماران داده شد.
موارد بكار رفته در تهیه دارو شامل تركیبات زیر بود: تریاك یا اپیوم خام تهیه شده از ستاد مبارزه با موادمخدر طی نامه شماره 1875 مورخ 15/1/78، اوسرین، وازلین، پروپیلن گلیكول، اتانول 96 درجه، متیل پارابن، پروپیلن پارابن، محلول سود 5%
تریاك حاوی آلكالوئیدها و مواد دیگری است كه بخشی از آن محلول در آب و بخشی محلول در الكل است. در ابتدا تریاك در الكل حل گردید. سپس مقدار بسیار مختصری سود 5% اضافه گردید و از صافی عبور داده شد. بدین ترتیب عصارة الكلی تریاك تهیه گردید. سپس مواد باقیمانده در آب حل گردید و مجدداً مختصری سود اضافه گردید و از صافی عبور داده شد و عصارة محلول در آب تریاك جدا گردید. كاربرد سود باعث تسهیل در جداسازی الكالوئیدهای تریاك میگردد. بدین ترتیب عصارة الكلی، عصارة محلول در آب و مواد باقیمانده بدست آمد. اوسرین و وازلین در 120 درجه سانتی گراد به مدت یك ساعت حرارت داده شد. سپس عصارة محلول در آب با اوسرین و عصاره الكلی و مواد باقیمانده با وازلین مخلوط گردید. از پروپیلن گلیكول بعنوان عامل افزایش دهنده نفوذ و جاذب رطوبت،متیل پارابن و پروپیل پارابن بعنوان محافظ استفاده گردید.
بدین ترتیب تركیبات دارویی زیر تهیه شد:
1-عصارة الكلی تریاك در وازلین 10%؛
2-عصارة محلول در آب تریاك در اوسرین 10%؛
3-مواد باقیمانده در وازلین 10%.
برای تهیه دارونما، از مواد ذكر شده بدون تریاك استفاده گردید و برای ایجاد رنگ مشابه، Cochineal red بعنوان ماده رنگی به كار رفت.
تمامی مراحل تولید دارو تحت شرایط آسپتیك انجام گرفت و فرآوردهها در تیوبهای مخصوصی كه از قبل حرارت داده شده بود جای داده شدند و روی تیوبها، كدهای مخصوصی نوشته شد. تمامی مراحل تولید و كدگذاری دارو در آزمایشگاه دانشكده داروسازی تحت نظر متخصص داروساز صنعتی بوده است.
مشخصات بیماران تحت مطالعه در پرسشنامة مخصوصی یادداشت و نتیجه معاینات بالینی ثبت و از بیماران یا والدین رضایتنامه كتبی اخذ میگردید. به هر بیمار یك ظرف دارو كه كد مخصوصی داشت تحویل میگردید و در پرسشنامه كد دارو نوشته میشد. به بیماران توصیه گردید كه قبل از استفاده از دارو، ضایعه را با آب معمولی شسته و خشك نمایند و بعد از استعمال دارو، روی آن را با گاز استریل بپوشانند. دارو دوبار در روز و به مدت دو هفته استعمال میگردید. بیماران جهت پیگیری هفتگی مراجعه میكردند . تأكید گردید در صورت بروز مشكل، سریعاً به پزشك مربوطه مراجعه نمایند.
در پایان هفته دوم، نتیجه معاینات بالینی و عوارض مشاهده شده ثبت میگردید. در بیمارانیكه پاسخ خوبی به درمان داده بودند در پایان هفته دوم مجدداً آزمایش مستقیم از ضایعه بعمل میآمد و در صورت منفی بودن آزمایش، به بیمار توصیه میگردیدكه هر دو هفته یكبار مراجعه و ضایعه مورد ارزیابی قرار گیرد. در پایان هفته هشتم، مجدداً از ضایعه آزمایش مستقیم به عمل میآمد و در صورت منفی بودن آزمایش و بهبود بالینی ضایعات، بیمار به عنوان درمان شده در نظر گرفته میشد.
به طور كلی معیارهای ارزیابی بیماران عبارت بودند از:
1-بهبودی كامل یا درمان شده: كاهش قابل توجه در اندازه، سفتی و التهاب ضایعه به همراه آزمایش مستقیم منفی ا زضایعه.
2-بهبود نسبی: كاهش قابل توجه در اندازه، سفتی و التهاب ضایعه ولی آزمایش مستقیم مثبت یا كاهش نسبی در اندازه، سفتی و التهاب ضایعه با آزمایش مستقیم منفی.
3-عدم پاسخ: عدم بهبودی ضایعه.
بیمارانی كه پاسخ نسبی یا عدم پاسخ به درمان دادند روی درمان دیگری قرار میگرفتند. در پایان مطالعه كدهای ثبت شده در پرونده با نوع داروی مصرفی تطبیق داده شد. اطلاعات ثبت شده در پرونده بیماران جمع آوری گردید و نتایج به دست آمده مورد تجزیه و تحلیل قرار میگرفت. در پایان مطالعه نوع داروی مصرفی با توجه به كدهای ثبت شده در پرونده مشخص گردید و اطلاعات موجود در پروندهها جمعآوری گردید و نتایج به دست آمده مورد تجزیه و تحلیل با آزمون آماری chi-square قرار گرفت.
یافتهها
در این مطالعه 96 بیمار (58 مرد و 38 زن) تحت درمان قرار گرفتند كه از این تعداد 52 بیمار دارو و 44 بیمار دارونما دریافت نمودند. در گروه دریافتكنندة دارو از 52 بیمار، 34 بیمار و از گروه دریافت كننده دارونما از 44 بیمار، 24 بیمار دورة درمان را به پایان رساندند و در مطالعه قرار گرفتند. بدین ترتیب 58 بیمار (37مرد و 21 زن) تحت درمان قرار گرفتند و دوره درمان را به پایان رساندند و 38 بیمار (21 مرد و 17 زن) از مطالعه خارج شدند.
گروه دریافت كننده دارو شامل 34 بیمار (21 مرد و 13 زن) بوده كه سن متوسط آنها 2/18 سال و تعداد ضایعات بطور متوسط 2 عدد و شایعترین شكل بالینی ضایعات بصورت پلاك زخمی بود. شایعترین محل ضایعه اندام فوقانی و طول متوسط شروع بیماری 2/2 ماه بود. هیچكدام از بیمارانیكه دارو را استعمال كردند عارضه جانبی خاصی را ذكر نكردند. در این گروه، 5 بیمار (7/14%) پاسخ كامل، 3 بیمار (9/8%) پاسخ نسبی و 26 بیمار(4/76%) به درمان پاسخ ندادند.
گروه دریافت كننده دارونما شامل 24 بیمار (16 مرد و 8زن) بوده كه سن متوسط آنها 4/21 سال بود. تعداد ضایعات بطور متوسط 6/2 عدد و شایعترین شكل بالینی ضایعات بصورت پلاك زخمی بود. شایعترین محل ضایعه اندام تحتانی و طول متوسط شروع بیماری1/2 ماه بود. در این گروه، 1 بیمار (2/4%) پاسخ كامل، 2 بیمار(3/8%) پاسخ نسبی و 21 بیمار (5/87%) به درمان پاسخ ندادند.
با استفاده از تست آماری chi-square ، از نظر آماری اختلاف معنیداری درپاسخ به درمان بین دو گروه وجود نداشت(12/0=P).
بحث
تاكنون اقدامات درمانی مختلفی در درمان سالك بكار رفته است كه بطوركلی شامل درمان فیزیكی یا جراحی و درمان دارویی میباشد. درمان فیزیكی و جراحی شامل كرایوتراپی، گرمای موضعی، كورتاژ و لیزرآرگون میباشد(9-6). درمان دارویی شامل درمان سیستمیك و موضعی است. مهمترین داروهای سیستمیك عبارتند از : آنتی موانهای پنج ظرفیتی (گلوكانتیم و پنتوستام)، كلروكین، پنتامیدین، مترونیدازول، كتوكونازول، داپسون، ایتراكونازول، تربینافین و ریفامپیسین(10).
داروهای موضعی به كار رفته شامل: كیناكرین، مایكونازول، كلوتریمازول، گلوكانتیم، كلروپرومازین، پارامومایسین، آمفوتریسین، كرم سیر و دارویZH-E میباشند(13-10و 3 و 1).
تریاك یا اپیوم خام از شیره گیاه خشخاش با نام علمی Papavera Somniferum استحصال میگردد. تریاك حاوی 30 آلكالوئید (14) و مواد دیگری نظیر اسید مكونیك، مكونین، پورفیروزین، موسیلاژ، پكتین، مواد آلبومینوئید ، موم، كائوچو، رزین، املاح معدنی و مواد قندی است(4). مهمترین آلكالوئیدهای تریاك شامل مورفین، كدئین، تبائین، نوسكاپین، نارسیئن و پاپاورین هستند(14). مكانیزم اثر اپیوم از طریق گیرندههای آنها در مغز، طناب نخاعی و جاهای دیگر است. گیرندههای اپیوم مو (μ ) ، كاپا (κ ) و دلتا(δ ) میباشند كه همگی آنها درك درد را كاهش میدهند(15).
اپیوئیدها رویسیستمایمنیاثرات مختلفیدارند.آنها قادر به رهایی هیستامین از ماست سلها هستند و منجر به خارش وواكنشهایآلرژیك میشوند. همچنین روی لكوسیتها اثر داشته وممكنست سبب تغییراتی در سیستم ایمنی شوند(15). لنفوسیتها و ماكروفاژها دارای گیرندههایی هستند كه قادر به پاسخگویی به اندورفینها و انكفالینها (اپیوئیدهای اندوژن) میباشند. بعلاوه قادر به تولید مواد شبه اندورفین هستند كه عمل لنفوسیتها را تنظیم مینمایند(16).
اپیوم در طب سنتی، در موارد مختلفی استفاده میگردد. مهمترین كاربرد آن بعنوان ضد درد قوی است. موارد استفاده دیگر اپیوم در درمان اسهال ساده، سرفه، دیابت(درگذشته)، بیخوابی، دردها و هیجانات عصبی، صرع و حالات اضطرابی میباشد. ولی همیشه بایستی در نظر داشت كه استفاده اپیوم موجب وابستگی میگردد(4).
در بررسی اپیوم خام در شرایط آزمایشگاهی بر روی شكل تاژك دار انگل (پروماستیگوت) مشاهده گردید كه حداقل غلظت مهاری تحریك و رشد 50% انگل به ترتیب 1/1 و 45/0 میلیگرم در میلی لیتر و حداقل غلظت مهاری تحریك و رشد 100% انگل به ترتیب 4 و 2 میلی گرم در میلی لیتر بود. در بررسی اپیوم خام بر روی موشهای Balb/c كه بصورت تجربی آلوده به لیشمانیا گردیدند، بعد از دو هفته درمان با اپیوم موضعی 5/87% بهبود بالینی و 7/68% بهبود بالینی و آزمایشگاهی گزارش گردید(5).
در این مطالعه كه با استفاده از اپیوم موضعی به مدت دو هفته در درمان سالك انجام گرفت مشاهده گردید كه اختلاف معنی داری در پاسخ به درمان، بین گروه گیرنده دارو و دارونما وجود ندارد (12/0 = P). علت عدم تأثیر دارو میتواند دورة كوتاه درمان، ضعیف بودن غلظت تریاك در پماد، عدم نفوذ كافی دارو در ضایعه و یا حجم كم نمونههای مورد مطالعه باشد. همچنین این احتمال وجود دارد كه سالك در موش سیر بالینی متفاوتی باانسان داشته باشد. همچنین احتمال سوگرایی(Bias) در مطالعه قبلی تأثیر اپیوم موضعی در لیشمانیوزیس موش وجود دارد. بنابراین ما پیشنهاد میكنیم كه بررسی دارو با غلظت و تركیبات متفاوت، ابتدا در حیوانات آزمایشگاهی انجام گیرد و در صورت مثبت بودن نتایج، مجدداً مطالعه روی موارد انسانی انجام پذیرد.
منابع
1-Dowlati Y. Cutaneous leishmaniasis: Clinical aspect. Clin Dermatol 1997; 14: 425-31.
2-اردهالی ص، رضایی ح، ندیم ا. انگل لیشمانیاو لیشمانیوزها. تهران: مركز نشر دانشگاهی، 1363: 161.
3- اصیلیان ع. لیشمانیوز جلدی (سالك). اصفهان: انتشارات دانشگاه علوم پزشكی اصفهان 1371 :40-38 و 55-54.
4-زرگری ع. گیاهان دارویی. تهران: انتشارات دانشگاه تهران، 1368: 130-128.
5-جلالی ع، مراغی ش، كوچك م. بررسی اثر اپیوم موضعی (شیره گیاه Papavera Somniferum ) بر روی لیشمانیا. پایان نامه جهت دریافت درجه دكترای داروسازی،دانشگاه علوم پزشكی اهواز، 1375: 56-52.
6-Al Gindan H, Kubba R. Cryosurgery in old
world cutaneous leishmaniasis. Br J Dermatol 1998; 118: 851-54.
7-Junid AJ. Treatment of cutaneous leishmaniasis with infrared heat. Int J Dermatol 1986; 25: 470-72.
8-Currine MA. Treatment of cutaneous leishmaniasis by currettage. Br Med J 1983; 287: 1105-6.
9-Rakcheev AP, Chistiakova IA. The succesful treatment of cutaneous leishmaniasis with an argon laser. Veston Dermatol Venerol 1989; 12: 53-55.
10-Lerner EA, Grevelink SA. Leishmaniasis. In: Arndt K A, LeBoil P E, Robinson JK, et al (eds). Cutaneous medicine and surgery. Philadelphia:W.B.Saunders, 1996:1163-70.
11-Vardy D, Baranholz Y C. Topical amphotericin B for cutaneous leishmaniasis. Arch Dermatol 1997; 135: 856-57.
12-غلامی ع، خامسی پور ع، مؤمنی ع و همكاران. اثر كرم حاوی سیر 5% در درمان سالك. فصلنامه بیماریهای پوست، 1379 : 3 :6-2.
13-Zerehsaz F, Salmanpour R, Handjani F. A double-blind randomized clinical trial of a topical herbal extract (ZHE) vs systemic meglumine antimonate for treatment of
cutaneous leishmaniasis in Iran. Int J Dermatol 1999; 38: 610-12.
14-Ewans WC. Trease and Evans pharmacognosy. Philadelphia: W.B. Saunders, 1996: 368-69.
15-Robert M, Julien M D. A Prime of drug action. New York: WH Freeman and Company, 1997: 285-90.
16-Terr AI, Dubey DD, Yunis EJ. Physiologic and enviromental influences on the immune system. In: Stites D P, Terr A I (eds). Basic and clinical immunology. Prantice Hall International Inc,1991:187-85.
مجلات نمايهشده در ISI
عناوين مجلات نمايه شده در بانك اطلاعاتي ISI Web of Science
بانك اطلاعاتي ISI Web of Science از 3 مجموعه زير تشكيل شده است:
1- مجموعه بانك Science Citation Index Expanded با عنوان اختصاري SCI-EXPANDED
2- مجموعه بانك Social Sciences Citation Index با عنوان اختصاري SSCI
3- مجموعه بانك Arts & Humanities Citation Index با عنوان اختصاري A&HCI
براي بررسي وجود يك عنوان مجله خاص در اين بانك و يا براي دستيابي به فهرست مجلات نمايه شده در اين بانك كافي است روي نام بانك مربوطه كليك كرده و در صفحه مربوطه گزينه مناسب را انتخاب كنيد.
لازم بهذكر است كه با توجه به موضوع، اكثر نشريات رشتههاي علوم پزشكي در لينك اول بايستي جستجو شوند.
مطالب ذیل را با اجازه دوستان عزیز از سایت بسیار کاربردی آزمایشگاهی ذیل برایتان قرار می دهم :http://azkadeh.com

در باره :مالاريا (About Malaria)
تعريف بيماري:
مالارياي انساني كه به نامهاي پالوديسم(Paludism) ، تب حارَه اي، تب نوبه، تب و لرز، تب متناوب و تب جنگل هم ناميده مي شود، يك بيماري عفوني خوني است كه توسط تك ياخته اي از جنس پلاسموديوم (Plasmodium) ايجاد و توسط پشه هاي جنس آنوفل(Anopheles) منتقل مي گردد.
علائم مالاريا (Paroxysm) :
علامت متداول بيماري تب شديدي است كه همراه با ساير علائم مربوطه (لرز ، عرق) براي مدت 5 تا 8 ساعت ادامه يافته و سپس هر 2 يا 3 روز يكبار باز مي گردد. هر يك از اين حملات، نيروي بيمار را به شدت تحليل برده و باعث كاهش شديد فعاليت هاي بيمار در طي دوران بيماري مي گردد.
علائم مالاريا (Paroxysm) :
علامت متداول بيماري تب شديدي است كه همراه با ساير علائم مربوطه (لرز ، عرق) براي مدت 5 تا 8 ساعت ادامه يافته و سپس هر 2 يا 3 روز يكبار باز مي گردد. هر يك از اين حملات، نيروي بيمار را به شدت تحليل برده و باعث كاهش شديد فعاليت هاي بيمار در طي دوران بيماري مي گردد. حملات اوليه مالارياي حاد حداقل 2 هفته يا بيشتر طول مي كشد و در اثر ابتلاهاي مكرر و همراه با پيشرفت بيماري ، كم خوني و بزرگي طحال حادث شده كه در اكثر بيماران در طي يك يا دو هفته بعد از حمله اوليه مي توان آنرا لمس نمود. در صورتيكه عامل ابتلا انگل گونه فالسيپارم باشد مي تواند منجر به گلومرولونفريت ، افزايش شديد پارازيتمي، پديده جداسازي مويرگي (Sequestration)، گيجي ، تشنج ، كاهش فشارخون ، ادم ريوي، علائم گوارشي (مثل استفراغ ، دردهاي شكمي ، اسهال ، خونريزي روده اي) ، سندرم نفروتيك، كولاپس گردش خون، سيانوز و مرگ شود.
دوره نهفتگي بيماري(Incubation Period) بطور معمول از 9 تا 30 روز متفاوت است و ممكن است تا چندين ماه طول بكشد. گونه هاي پلاسموديوم ويواكس در مناطق معين جغرافيايي دوره نهفته طولاني و بيشتر از 10 ماه دارند.
1 يا 2 روز قبل از شروع علائم تعداد كمي انگل در گردش خون ظاهر مي شوند سپس با افزايش تعداد انگلها ، علائم مختلف شروع به تظاهر مي كنند كه عبارتند از سردرد ، خستگي ، دردهاي مبهم در عضلات و استخوانها و مفاصل ، احساس لرز و تب كه بدليل شبيه بودن علائم با بيماريهاي ويروسي مثل سرماخوردگي و آنفلوانزا(Influenza) بخصوص در مناطق غير آندميك تشخيص را بر مبناي اين بيماريها مي گذارند. در طي چند روز آينده وقتي كه تعداد انگلها در خون به حد كافي رسيد علائم مالاريا بروز مي نمايد
علايم متداول مالاريا در بين مراجعين به Virtual Naval Hospital در آمريكا
ابتدا الگو هاي تب بصورت منظم نيست ولي بعدا در صورتي كه دوره هاي شيزوگوني همزمان شود رفته رفته اين الگو ها حالت منظمي را به خود مي گيرد كه به آنSynchronized مي گويند. بطور كلي يك حملة مالاريا شامل مراحل زير است:
1- مرحله سرما (Cold Stage ) : ناگهاني شروع مي شود. بيمار شديداً احساس كسالت نموده، دچار سردرد و دردهاي استخواني شده و تمام بدن با سرماي شديد و غير قابل تحملي به مدت 1 ساعت شروع به لرز مي نمايد.
2- مرحله گرما ( Hot Stage ) : با بالا رفتن دماي بدن به حدود 40 تا 41 درجه سانتيگراد لرز متوقف شده و تب شديدي بدن را فرا مي گيرد كه مدت 1 تا 2 ساعت اين وضعيت ادامه دارد . دورة تب بر حسب گونه هاي مختلف پلاسموديوم متفاوت و به صورت زير مي باشد.
- تب 3 به 1 : حملات تب يك روز درميان روي مي دهد. عامل آن پلاسموديوم اووال ، ويواكس و فالسيپارم ميباشد.
- تب 4 به 1 : حملات تب دو روز درميان و توسط پلاسموديوم مالاريه ايجاد مي گردد.
3- مرحله تعريق ( Sweating Stage ) : پس از گذشت 1 تا 2 ساعت از شروع تب ، بيمار شروع به عرق نمودن كرده كه در پايان اين مرحله و با گذشت 5 الي 8 ساعت از شروع تب ، درجه حرارت بدن به حد طبيعي باز مي گردد، بيمار خسته اما بدون علامت است و تا شروع مرحلة بعدي احساس آرامش مي كند.
روشهاي انتقال مالاريا :
شيوع و انتقال هر نوع مالاريا چه در مناطق اندميك و چه به صورت اپيدميك به عوامل بيشماري مثل نوع و سوش انگل ، ميزان ايمني انسانها ، عادات زيستي پشه ها ، عادات زيستي انسانها ، شرايط محيطي از قبيل درجه حرارت، رطوبت، ميزان بارندگي، نوع گياهان منطقه نوع و روشهاي مبارزه و . . . بستگي دارد كه اين عوامل را ميتوان در ارتباط با سه عامل شخص سالم و آلوده بعنوان گيرنده و دهندة بيماري ، انگل پلاسموديوم بعنوان عامل ايجاد كنندة بيماري و پشه آنوفل بعنوان انتقال دهندة بيماري طبقه بندي نمود. بطور كلي انتقال مالاريا از طرق زير مي تواند صورت بگيرد:
· انتقال طبيعي : انتقال از طريق نيش پشه آنوفل
· انتقال اكتسبي : انتقال از طريق تلقيح
انتقال طبيعي مالاريا (انتقال از طريق نيش پشه آنوفل):
عمده ترين راه انتقال بيماري مالاريا انتقال از طريق نيش پشه آنوفل ماده مي باشد كه در سال 1897 توسط رونالد راس ثابت شد. اين پشه ها در اكثر كشورهاي مناطق معتدله و حاره و هر جا كه محل زيست مناسبي براي آنها فراهم شود وجود دارند.
نحوة انتقال به اين صورت مي باشد كـه پشه هاي ماده در طي خونخواري از شخص آلــوده، انگل را بلعيده و انگل در بدن پشه پس از طي دوره و زمان خاصي مجدداَ به فرم آلوده كننده درآمده و همراه با محتويـات بزاق طــي خونخواري بعدي بـه افراد سالم جامعه منتقـل مي گـردد و اين چرخه تا جايي كه توسط عامل بخصوصي قطع نگردد ادامـه پيدا ميكند.
در اين چرخه براي آلوده شدن پشه ها، علاوه بر وجود ناقل مناسب وجود انسان مخزن ( (Human reservoir داراي مقدار لازم گامتوسيت در خون وي ضروري مي باشد. بنابراين انسانهايي كه به تازگي آلوده شده اند نمي توانند بيماري مالاريا را انتقال دهند.
انتقال اكتسابي ( انتقال از طريق تلقيح ):
در صورتيكه مالاريا غير از انتقال توسط نيش پشه آنوفل از طُرق ديگر منتقل شود به اين نوع مالاريا، مالارياي تلقيحي مي گويند. در سال 1884 انتقال مالاريا از طريق انتقال خون توسطGerhardet ثابت گرديد. در سال 1911 اولين مورد مالارياي ناشي از انتقال خون در آمريكا بوسيله Woolsey مشاهده شد.
در مناطقي كه انتقال مالاريا بوسيله پشه آنوفل انجام مي گيرد اهميت انتقال مالاريا از ساير راهها تحت الشعاع موارد انتقال عادي آن قرار ميگيرد، ولي در مناطقي كه انتقال طبيعي مالاريا بوسيله پشه آنوفل قطع شده است ولي حاملان انگل وجود دارند در صورتيكه عامل مالاريا پلاسموديوم فالسيپارم باشد و بموقع تشخيص داده و درمان نگردند، مي تواند به مرگ بيمار منجر شود.
مكانيزم عمل اين نوع انتقال به اين صورت است كه در افراد مبتلا به مالاريا در صورتيكه درمان كامل يا اساسي انجام نگرفته باشد پس از توسعه مصونيت نسبي (Premunition) ، تعداد انگل در خون به حداقل رسيده و مانع بروز علائم باليني مي شود و گاهي تعداد انگل در خون به حدي كم است كه در آزمايش ميكروسكپي معمولي نمونه خون نمي توان آنها را ديد ولي اگر خون اين افراد حامل انگل به افراد سالم از نظر بيماري تلقيح گردد، انگلهاي مرحله شيزوگوني خوني موجود در گلبول قرمز شروع به تكثير كرده و در گيرنده خون علائم مالاريا ظاهر مي گردد .
انواع پلاسموديوم هاي انساني در مالارياي ناشي از انتقال خون به ترتيب پلاسموديوم ويواكس، پلاسموديوم مالاريه ، پلاسموديوم فالسيپارم و پلاسموديوم اووال بوده اند كه در سالهاي اخير جاي خود را به پلاسموديوم مالاريه داده است، بنابراين در مالارياي ناشي از انتقال خون پلاسموديوم مالاريه نقش اصلي را دارد. مالارياي تلقيحي موارد زير را شامل مي گردد :
1. انتقال از طريق انتقال خون
2. انتقال از طريق پيوند اعضاء
3. انتقال از طريق مالاريا تراپي
4. انتقال از طريق هموتراپي
5. انتقال از طريق مادر به جنين
6. انتقال از طريق سرنگ آلوده
7. انتقال بصورت تصادفي ( آلودگي شغلي )

-----------------------------------------------------------------
تاريخچه مالاريا ( Malaria History )
مالاريا احتمالاَ بيماري قبل از پيدايش انسان است. منشاء احتمالي مالاريا بايد قاره آفريقا باشد و با مهاجرت انسان در طي دوران نوسنگي به اروپا و آسيا ( خاورميانه ، هند ، چين ) انتشار يافته است. انتشار اين بيماري به آمريكاي مركزي و جنوبي احتمالاَ از آسيا در هزارة اول ميلادي بوده است. نحوة ورود مالاريا به جزاير كارائيب هنوز نامشخص بوده و حدس بر اين است كه يا از طريق آمريكا بوده يا از طريق برده ها به آنجا برده شده است.
مالاريا يكي از قديمي ترين بيماري شناخته شده انساني بوده، يك نشانگان دوره اي كه با تب و لرز همراه مي باشد و احتمالاً توصيف مالاريا مي باشد در ادبيات قديمي چين ، سال 700 ق. م.، و در ورقهاي پاپيروس، سال 1570 ق. م.، شرح داده شده است. همچنين مالاريا در نوشته هاي قديمي به زبان سانسكريت، قرن 5 ق. م. در آسيا و چين شرح داده شده است.
بقراط(Hippocrates) پزشك يوناني در قرن پنجم قبل از ميلاد به رابطه تب هاي متناوب با مرداب ها اشاره نموده و علائم مالاريا نظير بزرگي طحال را شرح داده است. تب ها بوسيله سقراط(Socrates) فيلسوف يوناني، قرن 4 ق. م.، به انواع مداوم، روزانه، سه گانه و چهارگانه تقسيم شدند و عامل سه نوع آخر مالاريا دانسته شد. در بررسيهاي تاريخي علت شكست ارتش آتن در محاصره بندر سيسيليان سيراكوس، سال 413 ق. م.، و علت مرگ اسكندر مقدوني(Alexandre) در سن 32 سالگي، سال 323 ق. م.، را بيماري مالاريا مي دانند.
نام مالاريا از دو لغت ايتاليايي " Malo " به معني بد و "Aria " به معني هوا گرفته شده است. به دليل اينكه رم (Rome) در زمانهاي قديم شهري بوده كه توسط زمينهاي باتلاقي احاطه مي شده است و انتقال مالاريا در اطراف اين شهر را كه به سبب ايجاد محل مناسب براي رشد پشه ها فراهم شده بود ناشي از هواي فاسد بر روي آب راكد مي دانستند، لغت مالاريا به معني آب و هواي بد به اين بيماري اطلاق شد.
بدنبال تحقيقاتي كه در زمينه شناخت مالاريا در طي ساليان متمادي انجام شد سرانجام در سال 1880، پلاسموديوم مالاريه بوسيله لاوران (Laveran) شرح داده شد. در سال 1890 پلاسموديوم ويواكس توسط گراسي و فلتي (Grassi & Feletti) نامگذاري شد. در سال 1897 پلاسموديوم فالسيپارم توسط ولچ (Welch) و در سال 1922 پلاسموديوم اووال توسط استفنس (Stephens) توصيف شدند.
رونالد راس(Ronald Ross) بدنبال تحقيقاتي كه به پيشنهاد سرپاتريك مانسون در سكندرآباد و كلكته هندوستان انجام داد توانست در سال 1897 اسپوروگوني يك گونه پلاسموديوم بنام پلاسموديوم رليكتوم (Plasmodium Relictum) كه در پرندگان وجود داشت را در پشه كولكس كشف و گزارش دهد و به اين طريق توانست انتقال مالاريا توسط پشه ها را اثبات كند. اثباتي كه منجر به دريافت جايزه نوبل در سال 1902 گرديد ، گرچه ايدة انتقال مالاريا توسط پشه ها قبلاَ توسط ديگران بخصوص توسط پزشك رومي ، لانسيني، در سال 1717 و آلبرت كينگ در سال 1882 پيشنهاد شده بود.
در همان زمان گراسي (Grassi) و همكاران چرخه جنسي مشابهي را در پلاسموديوم فالسيپارم و در پشه آنوفل گزارش دادند و پشه آنوفل را بعنوان پشه ناقل شناسايي كردند. چرخه خارج اريتروسيتي (Exoerythrocytic cycle) مالاريا در انسان در سال 1948 بوسيله Shortt و همكاران شناسايي شد.
كلمه مالاريا طبق نوشته فرهنگ آكسفورد براي اولين بار در سال 1740 ميلادي توسط Horace Walpole به عنوان يك لغت انگليسي بكار برده شد و ظاهراَ در سال 1827 توسط John Callough عملاَ وارد فرهنگ زبان انگليسي گرديده است.

-----------------------------------------------------------
اهميت مالاريا
مالاريا يكي از علل عمدة همه گيري هاي بزرگ گذشته بوده است. اين همه گيري ها به شكل دوره اي به دنبال بارندگي هاي بيش از حد و سيل در نواحي خشك و يا در دشت هاي حاصلخيز رودخانه اي اتفاق افتاده و بدبختي حاصله از چنين بلاهايي را دو چندان كرده است.
همه گيري هاي مالاريا تلاش براي سكونت در نواحي جنگلي و آبياري زمينهاي خشك را خنثي نموده و جمعيت هاي بي خانمان و آوارگان را مصيبت زده نموده است. اين همه گيري ها به دنبال غارت و جنگ برخاسته و با توقف لشكر كشي ها گسترده شده ، و بازسازي هاي سريع زندگي روستايي و شهري را مختل نموده است.
گرچه امروزه ، در سايه تلاشهاي فراوان و زحمات بي دريغ كاركنان بهداشتي و در اثر توسعه نظام خدمات بهداشتي درماني ، بهبود كيفي و كمي مراقبت ها و امكانات تشخيصي و درماني و از همه مهمتر ارتقاء سطح آگاهيها و باورهاي بهداشتي مردم و رويكرد آنها به سوي رفتارهاي بهداشتي شاهد موفقيت هاي چشمگيري در زمينه كاهش موارد مالاريا در بسياري از كشورهای دنيا هستيم ، ولي وضعيت بيماري در برخي از كشورها ، سهولت و تكرر رفت و آمدها ، پنهان ماندن علائم ابتلا به عفونت در تعدادي از مبتلايان موجب شده است تا هميشه يك نگراني ضمني نسبت به خطر وقوع همه گيري مالاريا وجود داشته باشد. از آنجاييكه همه گيري هاي مالاريا علاوه بر ايجاد مشكلات بهداشتي، بيشتر توسعة اقتصادي را تحت تاثير قرار مي دهند می توان اهميت مالاريا از دو جنبه بهداشتي و اقتصادي بررسی نمود.
اهميت بهداشتي موضوع :
مالارياي انساني يك بيماري عفوني خوني است كه حدود 40 درصد جمعيت دنيا را تهديد مي كند. اين بيماري علاوه بر اينكه در طي حملات خود نيروي بيمار را به شدت تحليل برده و باعث كاهش شديد فعاليت هاي بيمار در طي دوران بيماري مي گردد ، موجب تضعيف بهداشت و رفاه عمومي در خانواده ها شده ، حيات كودكان را به مخاطره مي اندازد و باعث فرسودگي بيش از حد جامعه و منابع انساني كشورها مي شود.
در حال حاضر اين بيماري هنوز هم مهمترين مشكل بهداشتي دنيا است و حدود 107 كشور در دنيا بعنوان كشورهاي مالاريا خيز محسوب مي گردند. مرگ و مير ناشی از مالاريا بين 1/1 تا ۷/۲ ميليون نفر در سال گزارش شده است.
اهميت اقتصادي موضوع :
مالاريا با توجه به ميزان ابتلا و مرگ و مير در بين طبقات و گروههاي سني مختلف به خصوص طبقات فعال و توليد كنندة يك كشور مثل كارگران و كشاورزان تاثير سويي برروي توليدات و اقتصاد آن كشور بخصوص در جوامع كشاورزي دارد. آثار اقتصادي مالاريا به خصوص در نواحي روستايي قابل ملاحظه است چرا كه اغلب بروز مالاريا در زماني از سال است كه با فصل كشاورزي مصادف است. زيانهاي اقتصادي وارده توسط مالاريا بطور خلاصه عبارتند از :
- زيانهاي اقتصادي حاصله از مرگ و مير جميعت فعال
- زيانهاي اقتصادي ناشي از كم شدن قدرت كار و ميزان توليد در اثر ضعف و كم خوني
- زيانهاي مربوط به هزينه هاي درماني و مراقبت هاي پزشكي
- زيانهاي حاصله از كم شدن و از دست دادن وقت به علت غيبت از كار
- كاهش محصول كشاورزي و صنعتي در اثر كاهش و كمبود حضور كارگران مربوطه
- زيانهاي حاصله ناشي از توام بودن بيماريهاي ديگر با مالاريا و منجر شدن به مرگ شخص
- زيان اقتصادي ناشي از عدم انجام كشاورزي و كشت و كار در مناطق مالارياخيز كه معمولا با داشتن آب فراوان بهترين مناطق كشاورزي به شمار مي رود

--------------------------------------------------------
مالاريا در ايران
مالاريا از قديم به عنوان يكي از مسائل بهداشتي مهم تلقي شده و هيج بيماري ديگري به اين اندازه موجب زيان هاي مالي و جاني جبران ناپذير در كشور نشده است. همان طور كه مشخص است ، مهم ترين استان هاي دارای مشكل در كشور، عبارتند از سيستان و بلوچستان, هرمزگان , كرمان و فارس كه در مجموع 96 درصد از كل موارد را بخود اختصاص مي دهند.
سیمای کلی مالاریا در کشور طی پنج سال اخیر (1381-1386)
تعداد موارد مالاریا در طی سالهای 81-86 به غير از سال 82 که به طور غیر منتظره ای نسبت به سال 81 ، افزایش یافته است، سير نزولی يکنواختی را نشان می دهد.
از عمده دلایل افزایش موارد در سال 82 می توان به بارندگی بی سابقه در ماههای مرداد و شهریور سال مورد نظر و به دنبال آن ایجاد ژیت های لاروی گسترده در مناطق شرقی استان هرمزگان و جنوب سیستان و بلوچستان ، کاهش درجه حرارت و در نتیجه ازدياد وفور ناقل، افزایش شدید بیماری در مناطق مالاریا خیز کشورهای هم جوار ، عملیات نامناسب کنترل ناقلین به دلایل مختلف تدارکاتی از جمله فرسودگی تجهیزات سمپاشی و کمبود وسایل نقلیه عنوان کرد. با رفع نسبی نقائص و اقدامات مناسب پیشگیرانه در دانشگاه های علوم پزشکی مناطق جنوب شرقی کشور و هوشیاری سیستم بهداشتی در سال 83 وضعیت کنترل بیماری بهبود یافته و کاهش پنجاه درصدی موارد مالاریا حاصل گردید. در سال86 به دنبال سیل و طوفان گونو و علی رغم پیش بینی های کارشناسان مبنی بر افزایش چند برابری موارد مالاریا ، خوشبختانه بدلیل عملکرد صحیح و اقدامات به موقع کارکنان شاغل در امر کنترل مالاریای استانهای جنوب شرقی تعداد موارد مثبت نسبت به سال 85 تغییر معنی داری نداشت.
در طی سالهای 81 تا 86 همچون سالهای گذشته بیشترین میزان آلودگی به مالاریای ویواکس تعلق داشته که از سال 81 با یک روند صعودی روبرو بوده است. بدیهی است این روند صعودی به استثنای سال 82 نشان دهنده انتقال محلی در مناطق مالاریا خیز جنوب و جنوب شرقی کشور می باشد. کاهش موارد فالسیپارم نيز به شکلی موٌید افزایش انتقال محلی می باشد. می توان چنین قضاوت نمود که با کاهش تعداد مهاجران افغانی و پاکستانی درصد آلودگی به فالسیپارم در حال کاهش می باشد و به همان میزان موارد ویواکس جایگزین آن می شود.
طی شش سال اخیر چهره مالاریا از نظر توزيع بيماری به تفکيک مليت در کشور تغییر داشته است، به طوریکه در سال 81 بیش از نیمی از بیماران کشور را افراد غیر ایرانی تشکیل می دادند، در حالیکه در حال حاضر تنها 15% موارد بیماری به افراد غیر ایرانی تعلق دارد. در طی این سالها به تدریج با خروج افاغنه از کشور روز به روز بر نسبت مبتلایان ایرانی افزوده شده است.
چهره بیماری به غیر از گروه سنی 14-5 سال که روند افزایشی را نشان می دهد. درگروه های سنی زیر 4 سال و بالای 15 سال کاهش قابل توجهی را نشان می دهد. علت کاهش موارد ابتلای بالای 15 سال را می توان به دلیل کاهش ورود مهاجرین جویای کار از کشور های همجوار دانست. به دلیل آنکه اکثر مهاجرین به کشور در سن بالای 15 سال و نیروی کار محسوب می شوند، طبیعی است که با خروج آنان از کشور چهره بیماری تغییر یابد.
سیمای بیماری در استان هرمزگان از الگوی کشوری تبعیت نمیکند به گونه ای که درصد ابتلای گروه سنی زیر 4 سال به طور فزاینده ای از سال 81 تا 86 ازدیاد یافته و به همان نسبت درصد ابتلا در دو گروه سنی 14- 5 سال و بالای سال 15 نيزکاهش یافته است. می توان اینگونه تفسیر کرد که اکثر موارد بیماری در مناطق مشکل دار این استان به ويژه منطقه بشاگرد در حد فاصل شهرستان های جاسک و ميناب حاصل انتقال محلی مالاریا است.
حدود 12% از نسبت بیماران مرد در طی این سالها کاسته شده است، که با توجه به کاهش مهاجرین افاغنه از تعداد کاسته شدن از تعداد مبتلایان مرد طبیعی به نظر می رسد.
روند بیماری به لحاظ محل سکونت نيز طی سالهای مزبور تغییر یافته و بیشتر به سمت روستا معطوف گشته و بر این اساس بار دیگر می توان نتیجه گرفت که بیماری مالاریا در ایران بیشتر چهره محلی یافته است. از آنجائیکه نیروی کار مهاجر بیشتر در شهرها و حاشیه آن متراکم هستند، در گذشته در شهرها موارد بیشتری از بيماری مشاهده می شد. از طرف دیگر بهبود وضعیت زندگی در شهرها و ارتقاء شاخص های رفاهی و نیز صنعتی شدن شهر ها، کمک قابل توجهی به جلوگیری از انتقال بیماری در مناطق شهری نموده است.
بررسی کاهش موارد وارده و انتقال از وارده در کنار افزایش انتقال محلی نشان می دهد که در حال حاضر موارد اتوکتنوئوس (Autochthonous) مالاریا (جمع موارد انتقال محلی و عود) در کشور در حال افزایش است و ضرورت برنامه ريزی بر مبنای طبقه بندی صحيح و شناسایی دقيق کانونها و اجرای عمليات کنترلی متناسب با طبقه بندی مناسب را بيش از پيش آشکار می نمايد.
رفرنس: برنامه حذف مالاریا درجمهوری اسلامی ایران (افق1400 )، وزارت بهداشت درمان و آموزش پزشکی - معاونت سلامت

--------------------------------------------------------
مشخصات و مناطق انتشار آنوفلهاي مهم دنيا
1- An. aconitus
مناطق انتشار: شرق هندوستان، سريلانكا، مالزي، هندوچين ، اندونزي، جنوب چين
نوع لانه لاروي: لانه هاي لاروي آفتابي، باتلاق ها، آبگيرها و نهرهاي داراي گياه، مزارع برنج، مجاري آبياري
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : اماكن داخلي و خارجي
خونخواري از روي: انسان، حيوانات اهلي.
2- An. albimanus
مناطق انتشار: آمريكا ( ايالت تگزاس و فلوريدا )، كلمبيا، اكوادور، ونزوئلا
نوع لانه لاروي: آب شيرين يا شور، مرداب هاي داراي گياه، لانه هاي لاروي آفتابي
محل خونخواري بالغ : اماكن داخلي و خارجي
استراحت بعد از خونخواري: داخل اماكن و ساختمانها
خونخواري از روي: انسان و حيوانات اهلي
3- An. albitarsis
مناطق انتشار: آمريكاي مركزي به طرف آمريكاي جنوبي، ترينيداد
نوع لانه لاروي: آبگيرها، باتلاق هاي بزرگ داراي جلبك، استخرهاي آفتابي
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري: در محيط خارجي
خونخواري از روي: انسان و حيوانات اهلي
4- An. aquasalis
مناطق انتشار: جزاير آنتيل، ترينيداد، توباكو، جزاير مجاور آمريكاي مركزي تا مناطق شمالي برزيل
نوع لانه لاروي : مرداب ها، باتلاق هاي آب شور و داراي جذر و مد ، خورها، رودخانه هاي آب شور
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : اغلب در محيط خارجي
خونخواري از روي : انسان و حيوانات اهلي
5- An. arabiensis
توضيحات: يكي از اسپس هاي آنوفل گامبيه، دومين ناقل مالاريا در دنيا
مناطق انتشار: اكثر كشورهاي آفريقايي بخصوص مناطق خشك صحرايي
نوع لانه لاروي: لانه هاي لاروي موقت مثل آبگيرها، جاي پاي حيوانات، مزارع برنج، چاله ها آب
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : در داخل و خارج اماكن
خونخواري از روي: انسان، حيوان
6- An. azetecus :
مناطق انتشار : مكزيك ، در ارتفاعات بين 2200 - 1500 متر از سطح دريا
نوع لانه لاروي : آبگيرها، درياچه هاي داراي گياه، استخرها و حوض ها، كانال ها حتي با آب آلوده
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : در محيط خارج
خونخواري از روي : انسان
7- An. balabacensis
توضيحات: ناقل مالاريا در ، سوماترا و ساراواك، گونه جنگلي و وحشي
مناطق انتشار: هند، برمه، هندوچين، تايلند، مالزي، تايوان، سريلانكا، فيليپين، جاوه، برونئو، چين.
نوع لانه لاروي: لانه هاي لاروي سايه دار، آبگيرها ي خاكي، جاي چرخ وسايل نقليه، جاي پاي حيوانات
محل خونخواري بالغ: اماكن خارجي
استراحت بعد از خونخواري : اماكن خارجي
خونخواري از روي: انسان، حيوانات
8- An. bellator
مناطق انتشار: ونزوئلا، سورينام ، برزيل ، گينه و ترينيداد
نوع لانه لاروي: آبهاي جمع شده در ميان برگهاي گياه Bromeliade ، لانه هاي لاروي نيمه سايه
محل خونخواري بالغ: هنگام روز در جنگل هاي سايه دار
استراحت بعد از خونخواري : در اماكن داخلي
خونخواري از روي : انسان و حيوانات اهلي ، ترجيحاَ انسان
9- An. campestris
توضيحات: ناقل مالاريا و فيلاريازيس ( فرم دوره اي شبانه W. buncrofti و B. malyi )
مناطق انتشار: تايلند و احتمالاَ مناطق جنوب شرقي آسيا، در طول سواحل مالايا
نوع لانه لاروي: لانه هاي لاروي نيمه آفتابي، آبهاي عميق داراي مقداري گياه، آبهاي شورمزه، مجاري آب، چاههاي خاكي، گوشه هاي سايه دار مزارع برنج
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : بيشتر در اماكن داخلي
خونخواري از روي: انسان، حيوانات.

10- An. claviger
توضيحات: فقط در خاورميانه ناقل است.
مناطق انتشار : انگلستان و بعضي از كشورهاي اروپايي تا شمال آفريقا، فلسطين، ايران، عراق، افغانستان
نوع لانه لاروي: استخرها، باتلاق ها، چاهها، آب انبارها، آبگيرهاي سنگي، لانه هاي آفتابي يا سايه دار
محل خونخواري بالغ : داخل و خارج اماكن انساني
استراحت بعد از خونخواري : داخل و خارج اماكن انساني
خونخواري از روي: انسان، حيوانات.
11- An. cruzi :
توضيحات: مهمترين ناقل مالاريا در مناطق ساحلي برزيل.
مناطق انتشار: كستاريكا، پرو، برزيل، پاناما، اكوادور، بوليوي، كلمبيا، ونزوئلا
نوع لانه لاروي: آبهاي جمع شده در ميان برگهاي گياه Bromeliade ، لانه هاي لاروي نيمه سايه
محل خونخواري بالغ و استراحت بعد از خونخواري : داخل و خارج اماكن
خونخواري از روي: انسان
12- An. culicifacies
توضيحات: ناقل مهم مالاريا در پاكستان، هند، بنگلادش، سريلانكا
مناطق انتشار: عمان، بحرين، ايران، عرا ق، افغانستان، پاكستان به طرف هند، سريلانكا، اندونزي ، برمه، تايلند
نوع لانه لاروي: انواع آبهاي صاف يا آلوده و گاهي شور، چاهها، باتلاق ها، مزارع برنج، لانه هاي نيمه سايه
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : غالباَ در داخل ساختمانها
خونخواري از روي: انسان، حيوانات اهلي را نسبت به انسان ترجيح مي دهد.
13- An. fluviatilis
توضيحات: ناقل مهم مالاريا در پاكستان، هند، بنگلادش
مناطق انتشار : عمان، بحرين، شرق عربستان سعودي، ايران، عرا ق، افغانستان، پاكستان، هند، سريلانكا، برمه ، اندونزي، تايلند، تايوان، ، چين
نوع لانه لاروي: غالباَ در آبهاي جاري از قبيل نهرهاي خاكي، آبگيرها، بستر رودخانه و مجاري آبياري
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : داخل و خارج اماكن
خونخواري از روي: انسان، حيوانات اهلي.
14- An. funestus
توضيحات: سومين ناقل مالاريا در دنيا، ناقل فيلاريازيس ( فرم دوره اي شبانه W. Buncrofti )
ناقل آربوويروسهاي Nyando و Tanga در شرق آفريقا
مناطق انتشار: تمام كشورهاي آفريقايي جنوب صحرا
نوع لانه لاروي: در آبهاي كم و بيش دائمي و داراي گياه و سايه دار، باتلاقها، كنارة نهرها، رودخانه ها
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : غالباَ در داخل اماكن، ولي در محيط خارجي هم استراحت مي كند
خونخواري از روي: عمدتاَ انسان، حيوانات اهلي را نيز نيش مي زند.
15- An. gambiae
توضيحات: مهمترين ناقل مالاريا در دنيا ، ناقل فيلاريازيس ( فرم دوره اي شبانه W. Buncrofti)
ناقل آربوويروسهاي Tataguine به انسان در غرب آفريقا
مناطق انتشار: تمام كشورهاي آفريقايي جنوب صحرا
نوع لانه لاروي: لانه هاي لاروي موقت مثل آبگيرها، جاي پاي حيوانات، مزارع برنج، چاله ها آب
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : غالباَ در داخل اماكن، ولي در محيط خارجي هم استراحت مي كند
خونخواري از روي: عمدتاَ انسان، حيوانات اهلي را نيز نيش مي زند.
16- An. labranchiae
توضيحات: در شمال آفريقا ناقل است.
مناطق انتشار : انگلستان و بعضي از كشورهاي اروپايي تا شمال آفريقا، مراكش، الجزاير، تونس
نوع لانه لاروي: آبهاي شور در باتلاق هاي سواحل، باتلاق ها، مزارع برنج، زه آب ها، نهرها
محل خونخواري بالغ : داخل اماكن انساني و حيواني و محيط خارجي
استراحت بعد ازخونخواري : داخل و خارج اماكن انساني و حيواني
خونخواري از روي: انسان، حيوانات اهلي.
17- An. letifer
توضيحات: ناقل مالاريا و فيلاريازيس ( فرم دوره اي شبانه buncrofti. W)
مناطق انتشار: هند، تايلند، مالزي، فيليپين، اندونزي، سوماترا، برونئو
نوع لانه لاروي: آبهاي راكد به ويژه در دشتهاي ساحلي، آبگيرها، باتلاق ها، لانه هاي لاروي سايه دار
محل خونخواري بالغ: غالباَ پناهگاههاي خارجي
استراحت بعد از خونخواري : غالباَ پناهگاههاي خارجي
خونخواري از روي: انسان، حيوانات
18- An. leucosphyrus
توضيحات: ناقل مالاريا در ، سوماترا و ساراواك، گونه جنگلي و وحشي
مناطق انتشار: مالايا، سوماترا، برونئو، ساراواك
نوع لانه لاروي: لانه هاي لاروي سايه دار، آبگيرها ي خاكي، جاي چرخ وسايل نقليه، جاي پاي حيوانات
محل خونخواري بالغ: اماكن خارجي
استراحت بعد از خونخواري : اماكن خارجي
خونخواري از روي: انسان، حيوانات.
19- An. lesteri
توضيحات: ناقل مهم مالاريا در برخي از مناطق چين، ناقل فيلاريازيس ( فرم دوره اي شبانه malayi. B)
مناطق انتشار: كره، تايلند، مالزي، تايوان، فيليپين، جاوه، برونئو، جنوب چين تا ژاپن
نوع لانه لاروي: آبهاي خنك و تميز، آبهاي شورمزه، لانه هاي لاروي سايه دار
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : بيشتر در اماكن داخلي
خونخواري از روي: انسان، حيوانات.

21- An. maculatus
توضيحات: ناقل مالاريا و فيلاريازيس ( فرم دوره اي شبانه W. buncrofti )
مناطق انتشار: هند، سريلانكا، تايوان، مالزي، فيليپين، اندونزي، برونئو، هندوچين، جنوب چين
نوع لانه لاروي: لانه هاي لاروي آفتابي ، زه آبها، آبگيرهاي حاشيه نهرهاي آب ، مزارع برنج،
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : غالباَ پناهگاههاي خارجي
خونخواري از روي: انسان، حيوانات اهلي.
22- An. melas
توضيحات: يكي از اسپس هاي آنوفل گامبيه، ناقل مهمي در بسياري از مناطق ساحلي آفريقا.
مناطق انتشار: مناطق ساحلي غرب آفريقا تا كنگو
نوع لانه لاروي: آبهاي شور ، مرداب ها و باتلاق هاي داراي درخت كرنا ( Mangrove )
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : غالباَ در داخل اماكن، ولي در محيط خارجي هم استراحت مي كند
خونخواري از روي: انسان، حيوانات.
23- An. merus
توضيحات: يكي از اسپس هاي آنوفل گامبيه، ناقل مهمي در بعضي از مناطق ساحلي شرق آفريقا.
مناطق انتشار: مناطق ساحلي شرق آفريقا
نوع لانه لاروي: آبهاي شور ، مرداب ها و باتلاق هاي سواحل
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : غالباَ در داخل اماكن، ولي در محيط خارجي هم استراحت مي كند
خونخواري از روي : انسان، حيوانات.
24- An. minimus
توضيحات: ناقل مالاريا و فيلاريازيس ( فرم دوره اي شبانه W. buncrofti )
مناطق انتشار: عمان، سريلانكا، هند، برمه، هندوچين، تايلند، مالزي، تايوان، سوماترا، جاوه، چين، فيليپين
نوع لانه لاروي: غالباَ در آبهاي جاري از قبيل نهرها، چشمه ها، مجاري آبياري، زه آبها، مزارع برنج
محل خونخواري بالغ: بيشتر در اماكن داخلي
استراحت بعد از خونخواري : بيشتر در اماكن داخلي
خونخواري از روي: انسان، حيوانات اهلي.
25- An. naruna
مناطق انتشار: هند، برمه، تايلند، سريلانكا، تايلند
نوع لانه لاروي: آبهاي راكد، آبگيرها، لانه هاي لاروي آفتابي يا سايه دار، جريان هاي ملايم آب، چاهها
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : بيشتر در اماكن داخلي
خونخواري از روي: انسان، حيوانات اهلي.
26- An. nigerrimus
مناطق انتشار: هند، سريلانكا، تايلند، مالزي، برمه، برونئو، هندوچين، چين
نوع لانه لاروي: لانه هاي لاروي آفتابي، مجاري آبياري، باتلاق هاي داراي گياه، استخرهاي عميق ، مزارع برنج،
محل خونخواري بالغ: غالباَ اماكن خارجي
استراحت بعد ازخونخواري: غالباَ پناهگاههاي خارجي
خونخواري از روي: انسان، حيوانات.
27- An. nuneztovari
توضيحات: ناقل مهمي در كشورهاي كلمبيا و ونزوئلا محسوب مي گردد.
مناطق انتشار: گينه، برزيل، بوليوي، كلمبيا، ونزوئلا
نوع لانه لاروي: آبهاي گل آلود، استخرهاي كوچك و آفتابي، آبگيرها، جاي پاي حيوانات
محل خونخواري بالغ: اماكن خارجي
استراحت بعد از خونخواري : در محيط خارجي
خونخواري از روي: حيوانات ولي انسان را هم در محيط خارجي نيش مي زند
28- An. pattoni
مناطق انتشار: چين بالاتر از 30 درجه عرض شمالي
نوع لانه لاروي: لانه هاي لاروي آفتابي، استخرهاي سنگي داراي آلگ، آبگيرهاي اطراف نهرهاي آب
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : اماكن داخلي و خارجي
خونخواري از روي: انسان
29- An. pharoensis
توضيحات: فقط در مصر ناقل مهم محسوب مي گردد.
مناطق انتشار : مصر و قسمت اعظم آفريقا و جنوب صحرا، فلسطين، عربستان سعودي، سوريه
نوع لانه لاروي: باتلاق ها، مزارع برنج، استخرها، استخرهاي داراي گياه
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : در محيط خارجي
خونخواري از روي: انسان، حيوانات.
30- An. pseudopunctipennis
مناطق انتشار: جزاير آنتيل، جنوب ايالات متحدة آمريكا تا آرژانتين
نوع لانه لاروي: آبگيرها، كنارة نهرهاي داراي گياه، لانه هاي آفتابي، گودالهاي آب باران، زه آبها
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري: در محيط خارجي
خونخواري از روي : انسان و حيوانات اهلي
31- An. punctimacula
مناطق انتشار : مكزيك ، به طرف آمريكاي مركزي تا پرو ، برزيل ، آرژانتين و ترينيداد
نوع لانه لاروي : آبگيرهاي كوچك، كنارة نهرهاي داراي گياه و سايه دار، باتلاق ها
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري: در محيط خارجي و داخلي
خونخواري از روي : انسان و حيوانات اهلي
32- An. sacharovi
توضيحات: ناقل مالاريا در فلسطين و كشورهاي خاورميانه ، بعنوان زيرگونه maculipennis An.
مناطق انتشار: ايتاليا، يونان، اروپاي شرقي، فلسطين، عربستان سعودي، سوريه، تركيه، ايران، عرا ق
نوع لانه لاروي: باتلاق ها، آبگيرها، آب هاي شيرين يا شور، استخرها ي داراي گياه و آفتابي
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : در داخل اماكن انساني و حيواني
خونخواري از روي: انسان، حيوانات.
33- An. sergentii
توضيحات: ناقل مالاريا در مصر، فلسطين، عربستان سعودي
انتشار: الجزيره، تونس، فلسطين، عربستان سعودي، سوريه، تركيه، ايران، عرا ق، مصر، افغانستان
نوع لانه لاروي: مزارع برنج كاري، زه آبها ، نهرهاي آب با جريان ملايم داراي آفتاب و نيمه آفتابي
محل خونخواري بالغ : داخل و خارج اماكن
استراحت بعد از خونخواري : در داخل اماكن و پناهگاههاي خارجي
خونخواري از روي: انسان، حيوانات.
34- An. sinensis
توضيحات: ناقل مالاريا و فيلاريازيس ( فرم دوره اي شبانه W.buncrofti و malayi.B )
مناطق انتشار: درياي مديترانه، هند، خاور دور
نوع لانه لاروي: آبهاي خنك و تميز، آبهاي شورمزه، لانه هاي لاروي سايه دار
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : بيشتر در اماكن داخلي
خونخواري از روي: انسان، حيوانات.
35- An. stephensi
توضيحات: ناقل مالاريا در مناطق انتشار خود و شهرها و حومة آنها
انتشار: عربستان سعودي، عمان، ايران، عرا ق، افغانستان، بحرين، پاكستان، سريلانكا، هند، برمه، چين
نوع لانه لاروي: آبهاي شيرين يا شور، چاهها، مجاري آب، كناره هاي رودخانه ها، كانالهاي زهكشي
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : غالباَ در داخل ساختمانها
خونخواري از روي: انسان، حيوانات.
36- An. superpictus
انتشار: عربستان سعودي، ايران، عرا ق، افغانستان، اردن، پاكستان، تركيه، يونان، نواحي مديترانه
نوع لانه لاروي: آبهاي جاري، آبهاي كم عمق، بستر رودخانه هاي سنگلاخي، لانه هاي لاروي آفتابي
محل خونخواري بالغ: داخل و خارج اماكن
استراحت بعد از خونخواري : بيشتر در داخل خانه ها و اصطبل ها ، به نسبت كمتر در غارها
خونخواري از روي: انسان، حيوانات.
37- An. sundaicus
توضيحات: ناقل مالاريا و فيلاريازيس ( فرم دوره اي شبانه W.buncrofti )
مناطق انتشار: هند، برمه، هندوچين، تايلند، مالزي، تايوان، سوماترا، جاوه، برونئو، چين، اندونزي
نوع لانه لاروي: آبهاي شور يا شورمزه يا شيرين، مردابها، باتلاقها، آبگيرها، لانه هاي لاروي آفتابي
زه آبهاي مناطق ساحلي داراي آلگ هاي در حال فساد يا گياهان هرزآبي،
محل خونخواري بالغ: اماكن داخلي و خارجي
استراحت بعد از خونخواري : بيشتر در اماكن داخلي
خونخواري از روي: انسان، حيوانات اهلي.
38- An. subpictus
توضيحات: ناقل احتمالي مالاريا در جاوه و اندونزي
مناطق انتشار: ايران، پاكستان، هند، برمه، هندوچين، تايلند، مالزي، اندونزي، سريلانكا، جزاير جاوه، چين،
نوع لانه لاروي: آبگيرها ي خاكي نزديك خانه ها، مجاري فاضل آب، گودالهاي خاك برداري، آبهاي شورمزه
محل خونخواري بالغ : اماكن داخلي و خارجي
استراحت بعد از خونخواري : اماكن داخلي و خارجي
خونخواري از روي : انسان، حيوانات.

--------------------------------------------------------
اسامی ۲۳۳ گونه از آنوفل های دنیا
aconitus
albimanus
algeriensis
annandalei
annularis
apicimacula
apoci
aquasalis
arabiensis
ardensis
argenteolobatus
argyritarsis
argyropus
arnoulti
aruni
atroparvus (maculipennis atroparvus)
atropos
austenii
azaniae
azevedoi
aztecus
baezai
balabacensis
barberellus
barbirostris
barbumbrosus
bengalensis
berghei
bradleyi
braziliensis
brohieri
brohieri var. masseguini
brucei
brumpti
brunnipes
buxtoni
bwambae
caliginosus
cameroni
campestris
carnevalei
caroni
carteri
christyi
cinctus
cinereus
cinereus cinereus
cinereus hispaniola
claviger
concolor
confusus
coustani
crawfordi
cristipalpis
culicifacies
culicifacies adenensis
cydippis
dancalicus
darlingi
daudi
deemingi
demeilloni
distinctus
domicola
dthali
dureni
eiseni
erepens
erythraeus
ethiopicus
evansae
faini
farauti
flavicosta
fluviatilis
fontinalis
freeborni
freetownensis
funestus
fuscicolor
fuscivenosus
gambiae
garnhami
gibbinsi
greassei
grenieri
griveaudi
hamoni
hancocki
hargreavesi
harperi
hispaniola
hodgkini
hughi
hyrcanus
implexus
implexus niger
indefinitus
insulaeflorum
interruptus
jebudensis
karwari
keniensis
kingi
kochi
kosiensis
labranchiae (maculipennis labranchiae)
lacani
leesoni
lesteri
letabensis
letifer
listeri
lloreti
longipalpis
lounibosi
lovettae
machardyi
maculatus
maculipalpis
maculipennis (maculipennis maculipennis)
maliensis
marshallii
marteri
martinus (maculipennis martinus)
mascarensis
melanoon (maculipennis melanoon)
materi sogdianus
melas
merus
messeae (maculipennis messeae)
millecampsi
milloti
minimus
montanus
mogholensis
mortiauxi
moucheti
moucheti bervoetsi
moucheti moucheti
mousinhoi
multicolor
murphyi
namibiensis
natalensis
neivai
nigerrimus
nili
nimbus
nitidus
njombiensis
nuneztovari
obscurus
oswaldoi
paludis
parapunctipennis
parensis
pauliani
peditaeniatus
persiensis (maculipennis persiensis)
peryassui
peseudopictus
petragnani
peyrassui
pharoensis
philippinensis
plumbeus
pretoriensis
pseudopunctipennis
pulcherrimus
punctimacula
punctipennis
pursati
quadriannulatus
quadrimaculatus
radama
rageaui
ranci
rhodesiensis
rhodesiensis rhodesiensis
rhodesiensis rupicolus
riparis
rivulorum
rodhaini
roperi
roubaudi
ruarinus
rufipes
rufipes broussesi
rufipes rufipes
sacharovi
salbaii
schwetzi
separatus
sergentii
sergentii macmahoni
sergentii sergentii
seydeli
sinensis
smithii
somalicus
somaliensis
splendidus
squamosus
stephensi
subalpinus (maculipennis subalpinus)
subpictus
sundaicus
superpictus
swahilicus
symesi
tchekedii
tenebrosus
tessellatus
theileri
triannulatus
turkhudi
upemba
vagus
vaneedeni
vanhoofi
vernus
vestitipennis
vinckei
walravensi
wellcomei
wellcomei ugandae
wellcomei ungujae
wellcomei wellcomei
wellingtonianus
whartoni
wilsoni
ziemanni
--------------------------------------------------------


آيين نامه تاسيس و اداره آزمايشگاه هاي تشخيص طبي و آسيب شناسي
متن قانون:
ماده 1 تعريف :
1- آزمايشگاه تشخيص طبي و آسيب شناسي ، موسسه پزشكي است كه طبق ضوابط قانوني ايجاد مي گردد و در آن نمونه هاي حاصل از بدن انسان براي تشخيص و كنترل بيماريها و تاثير درمان مورد آزمايش قرار مي گيرد، هر آزمايشگاه تشخيص طبي نسبت به نوع تخصص و صلاحيت علمي مسئول يا مسئولين فني و براساس مجوزهاي صادره از وزارت بهداشت، درمان و آموزش پزشكي داراي يك يا چند بخش از بخشهاي زير مي تواند باشد.
الف) بيوشيمي
ب) خون شناسي
ج) ميكروب شناسي ( باكتري شناسي، انگل شناسي،ويروس شناسي ، قارچ شناسي)
د) ايمني شناسي و سرم شناسي
ه) آسيب شناسي تشريحي و ياخته شناسي (سيتولوژي و پاتولوژي)
و) ژنتيك پزشكي و مولكولي
ماده 2 تاسيس :
1-2) تاسيس آزمايشگاه قبل از هر گونه اقدام منوط به كسب موافقت اصولي و يا مجوز از وزارت بهداشت، درمان و آموزش پزشكي مي باشد.
2-2- اجازه تاسيس آزمايشگاه به كساني داده مي شود كه داراي شرايط ذيل باشند:
الف) تابعيت جمهوري اسلامي ايران
ب) نداشتن سوء پيشينه كيفري موثر
ج) دارا بودن يكي از مدارك تعيين شده ذيل:
دكتراي تخصصي آسيب شناسي باليني وتشريحي (يا هردو)
دكتراي تخصصي علوم آزمايشگاهي
دكتراي حرفه اي علوم آزمايشگاهي
دكتراي تخصصي تك رشته اي علوم آزمايشگاهي
تبصره 1- نهادها و موسسات دولتي و يا وابسته به دولت كه از بودجه عمومي استفاده مي نمايند به شرطي مي توانند تقاضاي تاسيس آزمايشگاه نمايند كه داراي مجوز قانوني، اجازه تاسيس موسسات پزشكي قيد باشد
تبصره 2- به موسس ازمايشگاه بيش از يك پروانه تاسيس داده نمي شود
تبصره 3- احراز صلاحيت موسس يا موسسين براساس شرايط فوق الذكر(ماده 2 و تبصره هاي مربوطه) به عهده كميسيون قانوني مصرح در ماده 20 مي باشد.
تبصره 4) در موارديكه موسس آزمايشگاه يك نفر باشد و شخص مزبور فوت نمايد وراث او مي توانند با ارائه گواهي تسليم حصر وراثت و معرفي يك نفر بعنوان مسئول فني واجد شرايط دريافت پروانه به وزارت بهداشت، درمان و آموزش پزشكي درخواست صدور پروانه مسئوليت فني موقت نمايند. اعتبار اين پروانه به مدت 2 سال خواهد بود. وراث مكلفند ظرف مهلت يادشده با ارائه دادنامه حصر وراثت نسبت به معرفي شخص و يا اشخاص واجد شرايط قانوني دريافت پروانه بعنوان موسس جديد اقدام كنند در غير اينصورت موسسه تعطيل خواهد شد.
د) انتقال پروانه تاسيس به غير توسط موسس ممنوع است مگر با موافقت كميسيون قانوني ماده 20 ضمناً براي افرادي كه يكبار پروانه تاسيس گرفته باشند و بهر علتي به شخص يا اشخاص واجد شرايط ديگري واگذار كرده باشند براي بار دوم پروانه تاسيس صادر نخواهد شد. ولي با رعايت ساير مقررات، پروانه تاسيس يك آزمايشگاه را مي توان به آنها منتقل نمود.
ر) درمانگاهها مي توانند با افزايش بخش آزمايشگاه به پروانه تاسيس مركز مذكور و با معرفي مسئول فني واجد شرايط به دانشگاه ها و موافقت كميسيون قانوني در اين رشته فعاليت نمايند.
ز) كميسيون قانوني با رعايت قوانين مصوب و مفاد اين آييننامه اقدام به بررسي و صدور پروانه خواهد بود. ماده 3 : انتقال بهره برداري، تعطيل 1-3) انتقال آزمايشگاه دائر از يك محل ديگر منوط به موافقت دانشگاه علوم پزشكي وخدمات بهداشتي درماني مقصد با اخذ پروانه تاسيس جداگانه طبق ضوابط مربوطه خواهد بود.
2-3) آزمايشگاه هاي بيمارستانهاي خصوصي مي بايستي در داخل محوطه مربوطه داير گردند و حق نصب تابلوي جداگانه بعنوان آزمايشگاه در محوطه خارج از بيمارستان را ندارند و فقط مي توانند نام آزمايشگاه، نام مسئول فني و تخصص مربوطه رادر تابلوي راهنماي بيمارستان قيد نمايند.
3-3) شروع بكار ، بهره برداري و ادامه فعاليت آزمايشگاه منوط به معرفي كاركنان پزشكي و پيراپزشكي و اعلام تجهيزات موردنياز و ساير مدارك طبق ضوابط اعلام شده توسط دانشگاههاي علوم پزشكي و خدمات بهداشتي درماني مربوطه و تاييد كميسيون قانوني آزمايشگاهها و كسب پروانه و رعايت ضوابط قانوني و مقررات مندرج در اين آييننامه و دستورالعمل هاي مربوطه خواهد بود.
4-3) تعطيل آزمايشگاه حداكثر تا شش ماه مجاز است مگر در مورد ماموريتهاي اداري، آموزشي و بيماري كه مورد قبول دانشگاه علوم پزشكي و خدمات بهداشتي درماني مربوطه قرار گيرد و در هر صورت موضوع تعطيلي آزمايشگاه بايد به اطلاع دانشگاه مربوطه برسد. چنانچه اين امر رعايت نگردد دانشگاه علوم پزشكي و خدمات بهداشتي درماني ذيربط مي تواند با توجه به نياز منطقه در محدوده آن آزمايشگاه به فرد ديگري طبق ضوابط و يا اعلام نظر نهايي توسط كميسيون قانوني آزمايشگاهها اجازه تاسيس آزمايشگاه بدهد. تبصره ) افراديكه بدون اطلاع اقدام به تعطيلي آزمايشگاه بيش از دو ماه بنمايند دانشگاه علوم پزشكي و خدمات بهداشتي درماني ذيربط، مي تواند با توجه به نياز منطقه در محدوده آن آزمايشگاه به فرد ديگري طبق ضوابط اجازه تاسيس آزمايشگاه بدهد.
ماده 4 : شرح وظايف موسس:
1-4) راه اندازي آزمايشگاه حداكثر ظرف مدت 2 سال پس از صدور موافقت اصولي
2-4) معرفي مسئول يا مسئولين فني آزمايشگاه به كميسيون قانوني
3-4) پاسخگويي در برابر مراجع قانوني در همه موارد بجز موارديكه به عهده مسئول فني بوده و در ماه 7 ذكر شده است.
4-4) تامين نيروي انساني واجد شرايط طبق نظر مسئول فني
5-4) اجرا و نظارت بر حفظ شئون اخلاق پزشكي و مقررات كشور
6-4) كنترل و نظارت بر حسن اجراي ضوابط، مقررات و تعرفه هاي مصوب
7-4) همكاري وايجاد تسهيلات لازم براي بازرسي نمايندگان وزارت بهداشت، درمان و آموزش پزشكي در هر زمان
8-4) ترتيب عمل دادن به توصيه ها و دستورات وزارت بهداشت، درمان و آموزش پزشكي در جهت حسن انجام امور آزمايشگاه
9-4) رعايت كليه قوانين و مقررات كشور درمورد نحوه اداره اماكن درخصوص نحوه بكارگيري افراد و نحوه پرداخت حقوق قانوني كاركنان
ماده 5- ساختمان آزمايشگاه
ساختمان آزمايشگاه بايد كليه ضوابط مربوط به شرايط فيزيكي يك آزمايشگاه تشخيص طبي را كه توسط وزارت بهداشت، درمان و آموزش پزشكي تعيين مي شود دارا باشد و در صورتي اجازه بهره برداري صادر مي شود كه بازرسين وزارت بهداشت، درمان و آموزش پزشكي اين شرايط را احراز نمايند. ماده 6- نيروهاي فني آزمايشگاه :
افراديكه در آزمايشگاه به كارفني مشغول مي شوند به شرح ذيل هستند:
1-6) مسئول فني آزمايشگاه (طبق ماده 7 اين آين نامه)
2-6) كارشناس ارشد باليني، يا كارشناس ارشد يكي از رشته هاي علوم آزمايشگاهي
3-6) كارشناس ازمايشگاه (ليسانس علوم آزمايشگاهي يا رشته هاي وابسته)
4-6) كاردان آزمايشگاه (فوق ديپلم)
5-6) تكنسين آزمايشگاه (ديپلم) كه دوره هاي آموزشي مربوطه را طي كرده باشد و يا پنج سال سابقه خدمت در آزمايشگاه راداشته باشد
تبصره 1) كليه افراد فوق بايد با صلاحديد و موافقت مسئول فني بكار گمارده شوند.
تبصره 2) كليه كادر فني بايد به دانشگاه مربوطه معرفي شوند.
ماده 7: مسئول فني
1-7) مسئول فني بايد داراي يكي از مدارك زير باشد:
دكتري تخصصي آسيب شناسي باليني يا تشريحي (يا هر دو)
دكتراي تخصصي علوم آزمايشگاهي
دكتراي حرفه اي علوم آزمايشگاهي
دكتراي تخصصي تك رشته اي علوم آزمايشگاهي
تبصره ) متخصصين تك رشته اي تنها مسئوليت فني آزمايشگاهي را مي توانند بپذيرند كه در همان رشته فعاليت مي نمايد. در صورتيكه متخصصان تك رشته اي بخواهند مسئوليت فني آزمايشگاه تشخيص طبي را بپذيرند لازم است چهار نفر رشته ، همزمان معرفي شوند.
2-7) مسئول فني توسط موسس يا موسسين به كميسيون قانوني معرفي مي شود و پس از احراز صلاحيت آنها و صدور پروانه مسئوليت فني مي تواند كار خود راشروع نمايد.
تبصره – موسس يا موسسين به شرط دارا بودن شرايط مسئوليت فني مي توانند به عنوان مسئول فني معرفي شوند
3-7) براساس مقتضيات زمان وزارت بهداشت، درمان و آموزش پزشكي مي تواند بطور موقت شرايط خاصي رابراي مسئوليت فني تعيين و ابلاغ نمايد.
4-7) هر مسئول فني مي تواند مسئوليت فني يك موسسه غيردولتي (اعم از خيريه، شركتها ، نهادها، خصوصي) را دريك نوبت كاري دارا باشد.
پروانه مسئول فني آزمايشگاههاي بيمارستانها و ساير مراكز درماني، به استناد پروانه تاسيس بيمارستان و يا درمانگاه مربوطه صادر خواهد شد.
5-7) صدور پروانه تاسيس مانع از پذيرش مسئوليت فني در آزمايشگاههاي تشخيص طبي براي موسس نخواهد بود.
ماده 8 شرح وظايف مسئول فني:
مسئول فني آزمايشگاه كسي است كه ضمن داشتن تخصص مربوطه، پروانه مسئوليت فني برابر قوانين و مقررات بنام او صادر شده ودخالت وي در امور آزمايشگاه فقط در حد صلاحيت مندرج در پروانه مسئوليت فني و دستورالعملهاي مربوطه آيين نامه مي باشد.
وظايف مسئول فني آزمايشگاه عبارت است از:
1-8- سرپرستي كليه امور تخصصي موسسه
2-8- كنترل و مراقبت وضعيت بهداشتي، تجهيزات و لوازم آزمايشگاهي مواد شيميايي، معرفها و تطبيق همه اين همه اين موارد با استانداردهاي اعلام شده از طرف وزارت بهداشت، درمان و آموزش پزشكي
3-8- ايجاد هماهنگي لازم بين واحدها و بخشهاي مختلف
4-8- كنترل عملكرد كيفي كاركنان فني
5-8- ممانعت از تحميل هزينه ها و خدمات غير ضروري به بيماران
6-8- رسيدگي به شكايات در مورد امور تخصصي و علمي آزمايشگاه و پاسخگويي به مراجع ذيربط نظير وزارت بهداشت، درمان و آموزش پزشكي ونظام پزشكي ، مراجع قضائي و غيره
7-8- جلوگيري از اعمال پزشكي غيرمجاز
8-8- جمع آوري اطلاعات و آمار مربوط به ارائه كمي و كيفي خدمات وارسال گزارشهاي مربوط به مراجع ذيربط به حسب ضرورت
9-8- رعايت قوانين و مقررات وزارت بهداشت، درمان و آموزش پزشكي
10-8- اقدام و نظارت بر پذيرش بيماران و ارائه خدمات اوليه به بيماران اورژانسي
11-8- نظارت بر اجراي دقيق برنامه كنترل كيفي داخلي و خارجي و انعكاس و نگهداري آنها در دفاتر مخصوص
12-8- حضور فعال در آزمايشگاه در ساعات مندرج در پروانه
13-8- امضاء ورقه جواب آزمايشها بادر نظر گرفتن كليه شرايط علمي و فني
ماده 9- شرايط ويژه مسئول فني:
1-9) حضور مسئول فني در موسسه در زمان انجام امور فني آزمايشگاه ضروري است و در صورتيكه به هر علت استثنائي مسئول فني نتواند شخصا در زمان تعيين شده در آزمايشگاه حضور داشته باشد، لازم است فردي واجد شرايط و صلاحيت را بصورت موقت به عنوان قائم مقام يا مسئول فني همكار معرفي نمايد. ولي بهرحال مسئوليت تمامي اقدامات رخ داده كماكان به عهده مسئول فني صاحب پروانه خواهد بود.
2-9) مسئول فني مي تواند در سال 2 ماه فرد واجد شرايط را به جانشيني خود بعنوان مسئول فني موقت به دانشگاههاي علوم پزشكي مربوطه معرفي نمايد و اين مدت تا 6 ماه قابل تمديد بوده و در موارد ذيل مدت آن افزايش مي يابد.
الف) ماموريتهاي آموزشي، فرصتهاي مطالعاتي و اداري كه به تاييد وزارت بهداشت، درمان و آموزش پزشكي رسيده باشد.
ب) بيماري خود و يا بستگان درجه يك كه مجبور به ترك شهر و يا شهرستان محل كار خود براي مدت زماني بيشتر از زمان ذكرشده در ماده 2-9 (با تاييد دانشگاه علوم پزشكي و خدمات بهداشتي درماني ذيربط)
تبصره 1- مسئول فني آزمايشگاه در غياب خود ضروريست فرد واجد شرايط را بعنوان جانشين معرفي نمايد.
تبصره 2- در صورتيكه مسئول فني آزمايشگاه كسي را معرفي ننمايد، موسس يا مسئول فني موسسه محق است كه فرد صاحب صلاحيت را جهت مسئوليت فني به دانشگاه مربوطه معرفي نمايد.
تبصره 3- در صورتيكه مسئول فني آزمايشگاه يك مركز درماني اعم ازبيمارستان يا درمانگاه در غياب خود تا 6 ماه اقدام به معرفي فرد واجد شرايط ننمايد، موسس و يا مسئول فني موسسه اختيار خواهد داشت تا نسبت به معرفي يك نفر واجد صلاحيت براي مسئوليت فني دائم به دانشگاه مربوطه اقدام تا كسب صلاحيت نمايد.
3-9) متخصيصني كه توسط مسئول فني آزمايشگاه بعنوان همكار نيمه وقت و يا تمام وقت آزمايشگاه معرفي و اجازه مربوطه رادريافت مي دارند تا زمان ادامه همكاري ذكر نام آنها روي تابلو و اوراق بلامانع است. اين افراد نيز موظف به حضور در آزمايشگاه در ساعات مقرر مي باشند، بديهي است امضاء ورقه هاي آزمايش صرفاً با امضاء مسئول فني واجد شرايط داراي پروانه معتبر است
4-9) مسئول فني يك آزمايشگاه مي تواند با رعايت همه ضوابط مربوط به مسئوليت فني، همكار يك آزمايشگاه ديگر باشد.
5-9) احكام مسئول فني موقت آزمايشگاه توسط اداره امور آزمايشگاههاي دانشگاهها براساس ضوابط و شرايط فوق الذكر صادر خواهد شد.
ماده 10- شرايط و ضوابط عمومي نحوه كار در آزمايشگاه تشخيص طبي
1-10- نام موسسه ، آدرس و تلفن و ساعات كار بايد روي ورقه آزمايشگاه منعكس شود ضروريست نام مسئول فني و شماره نظام پزشكي بر روي ورقه هاي آزمايشگاه قيد شود.
2-10- آزمايشگاه موظف به داشتن دفتر كل و گزارش رايانه اي بيماران جهت ثبت نام بيماران و مشخصات كامل آنها و نام پزشك درخواست كننده، نوع آزمايشهاي درخواستي، قيمت، شماره سريال قبض نوع بيمه و تاريخ مي باشد.
3-10- كليه دفاتر آزمايش بايد حداقل به مدت يكسال در آزمايشگاه بايگاني شود . هريك از بخشهاي آزمايشگاه بايد دفتر جداگانه اي جهت ثبت نتيجه هاي آزمايش بيماران داشته باشد.
4-10- آزمايشگاه موظف به صدور قبض رسيد در مقابل دريافت وجه از بيماران بوده ورونوشت و يا ته قبض آن رابايد به مدت شش ماه نگهداري نمايد.
5-10- استاده از هر گونه نام و عناوين روي تابلو و ورقه هاي آزمايشگاه به غير از آنچه در پروانه قيد شده است ممنوع مي باشد.
6-10- نمونه برداري در آزمايشگاه بايد طبق ضوابط علمي صورت گرفته و مفاد آيين نامه انطباق امور اداري و فني موسسات پزشكي با موازين شرع مقدس دقيقاً رعايت شود.
7-10- آزمايشگاه موظف است به تناسب امكانات علمي و فني خود آزمايش بپذيرد.
8-10- تعداد آزمايشگاه موردنياز در هر شهر و فاصله آنها از همديگر براساس جمعيت، وسعت، پراكندگي جمعيت و ساير موضوعات توسط گروه كارشناسي متشكل از نمايندگان اداره امور آزمايشگاههاي دانشگاههاي علوم پزشكي شهرستانها و هماهنگي اداره امور آزمايشگاهها خواهد شد.
تبصره 1) آزمايشگاههاي بيمارستاني از اين بند مستثني هستند.
9-10- ظرفيت بايد براساس كل پذيرش انجام گرفته و نبايد هيچگونه تفاوتي بين بيماران بيمه اي و غيربيمه اي باشد وتنها پذيرش بر مبناي حق تقدم مراجعين مي باشد.
تبصره : بيماران اورژانس مستثني از اين قاعده هستند.
10-10- هر آزمايشگاه موظف است نرخ تعرفه خدمات آزمايشگاهي رادر مكاني مناسب و در محلي كه در معرض ديد همگان باشد نصب نمايد.
11-10- چون اجراي كنترل كيفي (داخلي و خارجي) براي آزمايشگاههاي كشور جزء وظايف اوليه محسوب شده است لذا در اجراي اين هدف آزمايشگاهها موظف به اجراء برنامه كنترل كيفي و نيز شرايط و ضوابط تعيين شده در طرح استاندارد سازي آزمايشگاههاي تشخيص طبي مي باشند.
ماده 11- شرايط و ضوابط عمومي نحوه كار در آزمايشگاههاي آسيب شناسي تشريحي(هيستوپاتولوژي) 1-11- كليه نسوج برداشته و مايعات خارج شده از بدن بيماران بايد براي آزمايشهاي هيستوپاتولوژي و سيتولوژي به آزمايشگاههاي مجاز براي انجام آزمايشهاي لازم ارسال گردد.
2-11- آزمايشگاه آسيب شناسي تشريحي بايد جهت آزمايش نمونه ارسالي، اطلاعات لازم شامل درخواست پزشك معالج، نام ونام خانوادگي بيمار- نام پدر- سن- جنس- محل برداشت نمونه و تاريخ برداشت آن- اطلاعات اساسي از شكايات و علائم باليني و آزمايشگاهي، مشاهدات حين بيوپسي و يا عمل جراحي و تشخيص يا تشخيص هاي افتراقي را اخذ و سپس به آزمايش اقدام نمايد.
در مورد نمونه هاي مربوط به دستگاه توليد مثل بانوان، وضع زنانگي، تاريخ آخرين قاعدگي و قاعدگي ماقبل و استفاده از هورمون ها نيز بايستي ذكر شود.
3-11- مشخصات بيمار، نام و نام فاميل، نام پزشك معالج و محل برداشت نمونه بايد روي ظرف حاوي نمونه بطور جداگانه و با مسئوليت پزشك معالج نوشته شود و با ورقه درخواست آزمايش تطبيق نمايد.
تبصره : جراح و يا پزشك معالج موظف است كه تمامي نمونه را با ذكر تمام مشخصات نمونه و بيمار فقط براي يك آزمايشگاه ارسال دارد ونبايستي نمونه مورد آزمايش به قطعاتي تقسيم و براي آزمايشگاههاي متعدد فرستاده شود، در صورت كشف اين موضوع آزمايشگاه مي تواند از پذيرش آن خودداري نمايد.
در صورتيكه كشف اين موضوع پس از برش و طي مراحل مختلف تكنيكي و قبل از دادن جواب آسيب شناسي باشد. آسيب شناس مخير است كه بقيه نمونه و لامهاي تهيه شده را پس از دريافت هزينه هاي مربوطه به صاحب آن مسترد نمايد و يا مانند هربيوپسي ديگري آزمايش ميكروسكوپي انجام داده وجواب آنرا بدهد.
ماده 12- شرايط ويژه آزمايشگاههاي آسيب شناسي تشريحي و ياخته شناسي(سيتولوژي)
الف- مسئول فني اين آزمايشگاه ها بايد داراي درجه دكترا در پزشكي ودرجه تخصصي در پاتولوژي تشريحي باشد.
ب- جواب پاتولوژي و سيتولوژي بايد حداقل دردو نسخه گزارش شود كه نسخه اصلي به بيمار داده شده يابراي پزشك معالج ارسال مي گردد و نسخه دوم در آزمايشگاه بايگاني مي شود.
ج- كليه بلوكها بايد براي هميشه در بايگاني آزمايشگاه نگهداري شود.
د- كليه لامهاي پاتولوژي بايد حداقل بمدت 10 سال در آزمايشگاه نگهداري شوند.
هـ - كليه لامهاي سيتولوژي بايد حداقل بمدت 6 ماه در آزمايشگاه نگهداري شوند و موارد مثبت يا مشكوك بايستي براي هميشه در آزمايشگاه نگهداري شوند. كپي نتايج گزارش همراه بادرخواست آزمايشات پاتولوژي حداقل 2 سال نگهداري شوند.
ز- كليه نمونه ها بايد با شماره اي مشخص شوند و در دفتر انديكس ، روي نمونه، روي لامها، روي بلوكها و در برگه گزارش اين شماره منعكس شود.
ح- نام، فاميل، نام پدر، سن و جنس بيمار و نام پزشك معالج و نيز تاريخ صدور جواب، محل برداشت و نوع بافت مورد آزمايش بايد در ورقه گزارش شود.
تبصره – مسئوليت دادن اين اطلاعات و صحت آن بعهده پزشك معالج است.
ط- نوع فيكساتور مصرف شده بايد ذكر شود.
ي- شرح ماكروسكوپي در گزارش لازم است و بايد مشخص شود كه بافت تماما يا قسمتي از آن مورد بررسي قرار گرفته است.
ك- تشخيص نهايي بايد بطور مشخص در گزارش منعكس گردد.
ل- ذكر نام پاتولوژيست همراه با امضاي وي در گزارش الزامي است.
م- در صورتي كه پزشك معالج مايل به اخذ نظر مشورتي ديگري باشد با درخواست كتبي، آزمايشگاه موظف است لامهاي موردنظر و يا درصورت لزوم بلوكها را دراختيار پزشك معالج قرار دهد. آزمايشگاه مورد مشورت بايد لامهاي موردنظر را شماره گذاري نمايد و همانند نمونه هاي ديگر گزارش و نگهداري نمايد، ولي بلوكها حتما به آزمايشگاه اول برگشت داده شوند. در صورتيكه به علل تكنيكي تنها در يك لام ضايعه ديده شود لام مذكور پس از مشاوره به آزمايشگاه اوليه عودت داده شود.
ماده 13- ضوابط مربوط به لغو موقت دائم پروانه آزمايشگاه
موسس و يا موسسين آزمايشگاه موظفند الزامات موضوع اين آيين نامه و دستورالعملهاي ابلاغ شده توسط وزارت بهداشت،درمان وآموزش پزشكي را رعايت نمايند و مسئول فني و كاركنان ذيربط نيز مكلف به انجام آنها در حدود وظايف و صلاحيتهاي مربوطه هستند و بايد نواقص اعلام شده رادر مدت زمان تعيين شده برطرف سازند و در صورت تخلف بسته به نوع تخلف به نحو زير اقدام خواهد شد.
الف) تذكر شفاهي با قيد موضوع در صورتجلسه كارشناسي محل توسط دانشگاههاي علوم پزشكي وخدمات بهداشتي درماني ذيربط
ب) اخطار كتبي و درج در پرونده توسط دانشگاه علوم پزشكي و خدمات بهداشتي درماني ذيربط
ج) لغو موقت پروانه تاسيس و تعطيل آزمايشگاه بمدت يكسال به پيشنهاد دانشگاه علوم پزشكي و خدمات بهداشتي درماني با نظر مراجع قانوني ذيصلاح در استانها
د) لغو دائم پروانه تاسيس آزمايشگاه به پيشنهاد دانشگاه علوم پزشكي و خدمات بهداشتي درماني با تصويب معاونت درمان و نظر مراجع قضائي ذيصلاح
ماده 14- اين آيين نامه باستناد قانون مربوط به مقررات امورپزشكي و دارويي موادخوراك و آشاميدني مصوب 29/3/1334 و اصلاحيه سال 23/1/1367 و قانون تشكيلات وظايف وزارت بهداشت درمان و آموزش پزشكي مصوب سال 1367 در تاريخ 28/1/78 به تصويب رسيد و كليه آيين نامه ها و دستورالعملها و بخشنامه هاي مغاير با آن از تاريخ اجراء اين آيين نامه لغو مي گردد.
سيستم هاي آنزيمي R-M نوع II:
نامگذاري آنزيمهاي R-M
2- بعد از سه حرف مذكور حرف اول سويه يا گونه بصورت غير ايتاليك و يا اعداد عربي مي آيد مانند: EcoK بري Ecoli سويه K ، Hind براي H.influenza سويه d يا Sau 3A براي استافيلوكوك اورئوس 3A . عناصر خارج كروموزومي مانند ويروس يا پلاسميد به همين صورت متعاقب اسم مذكور آورده ميشود مانند: EcoRI و EcoPI.
3- متعاقب آن بدون در نظر گرفتن فاصله ، اعداد روماني (I,II,III,.....) براي تعيين آنزيم هاي مختلف كه توسط همان سويه توليد ميشوند مانند HindII و HindIII .
4- براي اختصاصي كردن R.ENase يا M.MTase حروف بزرگ R و M در جلو نشانه آنزيم قرار ميگيرد و با يك نقطه از آن فاصله ميگيرد مانند : R.EcoRI و M.EcoRI.
جدول شماره 6- كد اسيد هاي نوكلئيك جايگاه شناسايي آنزيم ها ي محدود گر
|
اسيد نوكلئيك |
كد |
اسيد نوكلئيك |
كد |
|
A, C or T |
H |
G or A |
R |
|
A, C or G |
V |
C orT |
Y |
|
C, G or T |
B |
A or T |
W |
|
A, G or T |
D |
A or C |
M |
|
G, A, T or C |
N |
G or T |
K |
|
|
|
C or G |
S |
|
Third position (3` end) |
Second position |
First position (5` end) | |||
|
G |
A |
C |
U | ||
|
U |
Cys |
Tyr |
Ser |
Phe |
U |
|
U |
Arg |
His |
Pro |
Leu |
C |
|
U |
Ser |
Asn |
Thr |
Ilu |
A |
|
U |
Gly |
Asp |
Ala |
Val |
G |
GUG - بندرت به عنوان رمز شروع كننده قرار ميگيرد.
جدول شماره 8- حروف رمز اسيد هاي آمينه
|
نام اسيد آمينه |
رمز يك حرفي |
رمز سه حرفي |
نام اسيد آمينه |
رمز يك خرفي |
رمز سه حرفي |
|
آلانين |
A |
Ala |
لوسين |
L |
Leu |
|
آرژنين |
R |
Arg |
ليزين |
K |
Lys |
|
آسپاراژين |
N |
Asn |
متيونين |
M |
Met |
|
آسپارتيك |
D |
Asp |
فنيل آلانين |
F |
Phe |
|
سيتئين |
C |
Cys |
پرولين |
P |
Pro |
|
گلوتامات |
E |
Glu |
سرين |
S |
Ser |
|
گلوتامين |
Q |
Gln |
تره اونين |
T |
Thr |
|
گليسين |
G |
Gly |
تريپتوفان |
W |
Trp |
|
هيستيدين |
H |
His |
تيروزين |
Y |
Tyr |
|
ايزولوسين |
I |
Ile |
والين |
V |
Val |
atI AGG!CCT
AatII GACGT!C
AccI GT!MKAC
AccII CG!CG
AccIII T!CCGGA
AcyI GR!CGYC
AflI G!GWCC
AflII C!TTAAG
AflIII A!CRYGT
AhaI CC!SGG
AhaII GR!CGYC
AhaIII TTT!AAA
AluI AG!CT
AlwI GGATCNNNN!
AlwI !NNNNNGATCC
AlwNI CAGNNN!CTG
AocI CC!TNAGG
AocII GDGCH!C
AosI TGC!GCA
AosII GR!CGYC
ApaI GGGCC!C
ApaLI G!TGCAC
ApyI CC!WGG
AquI C!YCGRG
AseI AT!TAAT
Asp700I GAANN!NNTTC
Asp718I G!GTACC
AspI GACN!NNGTC
AsuI G!GNCC
AsuII TT!CGAA
AvaI C!YCGRG
AvaII G!GWCC
AvaIII ATGCA!T
AvrI CYCGRG
AvrII C!CTAGG
AxyI CC!TNAGG
BalI TGG!CCA
BamHI G!GATCC
BanI G!GYRCC
BanII GRGCY!C
BanIII AT!CGAT
BbeI GGCGC!C
BbiII/AcyI GR!CGYC
BbvI GCAGCNNNNNNNN!
BbvI !NNNNNNNNNNNNGCTGC
BbvII GAAGACNN!
BbvII !NNNNNNGTCTTC
BcefI ACGGCNNNNNNNNNNNN!
BcefI !NNNNNNNNNNNNNGCCGT
BclI T!GATCA
BcnI CC!SGG
BglI GCCNNNN!NGGC
BglII A!GATCT
BinI !NNNNNGATCC
BinI GGATCNNNN
BsmAI GTCTC
BsmAI GAGAC
BsmI GAATGCN!
BsmI G!CATTC
Bsp1286I GDGCH!C
BspHI T!CATGA
BspMI ACCTGCNNNN!
BspMI !NNNNNNNNGCAGGT
BspMII T!CCGGA
BsrI ACTGGN!
BsrI C!CAGT
BstBI TT!CGAA
BstEII G!GTNACC
BstI G!GATCC
BstNI CC!WGG
BstPI G!GTNACC
BstUI CG!CG
BstXI CCANNNNN!NTGG
BstYI R!GATCY
Bsu36I CC!TNAGG
CcrI C!TCGAG
CfoI GCG!C
Cfr10I R!CCGGY
Cfr13I G!GNCC
CfrI Y!GGCCR
ClaI AT!CGAT
CviJI RG!CY
CvnI CC!TNAGG
DdeI C!TNAG
DpnI GA!TC
DraI TTT!AAA
DraII RG!GNCCY
DraIII CACNNN!GTG
DsaI C!CRYGG
EaeI Y!GGCCR
EagI C!GGCCG
EarI CTCTTC
EarI GAAGAG
EclXI C!GGCCG
Eco105I TAC!GTA
Eco31I GGTCTCN
Eco31I !NNNNNGAGACC
Eco47I G!GWCC
Eco47III AGC!GCT
Eco52I C!GGCCG
Eco57I CTGAAG
Eco57I CTTCAG
Eco81I CC!TNAGG
EcoNI CCTNN!NNNAGG
EcoO109I RG!GNCCY
EcoRI G!AATTC
EcoRII !CCWGG
EcoRV GAT!ATC
EcoT14I C!CWWGG
EcoT22I ATGCA!T
EcoT38I GRGCY!C
EheI GGC!GCC
EspI GC!TNAGC
FinI GTCCC
FinI GGGAC
Fnu4HI GC!NGC
FokI GGATGNNNNNNNNN!
FokI !NNNNNNNNNNNNNCATCC
FspI TGC!GCA
GdiII !NNNNNYGGCCG
GdiII CGGCCRN!
GsuI CTCCAG
GsuI CTGGAG
HaeI WGG!CCW
HaeII RGCGC!Y
HaeIII GG!CC
HapII C!CGG
HgaI GACGCNNNNN!
HgaI !NNNNNNNNNNGCGTC
HgiAI GWGCW!C
HgiEII ACCNNNNNNGGT
HhaI GCG!C
Hin1I GR!CGYC
HinP1I G!CGC
HincII GTY!RAC
HindIII A!AGCTT
HinfI G!ANTC
HpaI GTT!AAC
HpaII C!CGG
HphI GGTGANNNNNNNN!
HphI !NNNNNNNTCACC
KpnI GGTAC!C
Ksp632I CTCTTCN!
Ksp632I !NNNNGAAGAG
MaeI C!TAG
MaeII A!CGT
MaeIII !GTNAC
MboI !GATC
MboII GAAGANNNNNNNN!
MboII !NNNNNNNTCTTC
MfeI CAATTG
MflI R!GATCY
MluI A!CGCGT
MmeI TCCRAC
MmeI GTYGGA
MnlI CCTCNNNNNNN!
MnlI !NNNNNNNGAGG
MroI T!CCGGA
MseI T!TAA
MspI C!CGG
MstI TGC!GCA
MstII CC!TNAGG
MvaI CC!WGG
NaeI GCC!GGC
NarI GG!CGCC
NciI CC!SGG
NcoI C!CATGG
NdeI CA!TATG
NdeII !GATC
NheI G!CTAGC
NlaIII CATG!
NlaIV GGN!NCC
NotI GC!GGCCGC
NruI TCG!CGA
NsiI ATGCA!T
Nsp(7524)I RCATG!Y
Nsp(7524)V TT!CGAA
NspBII CMG!CKG
NspII GDGCH!C
NspIII C!YCGRG
NspIV G!GNCC
NunII GG!CGCC
PaeR71 C!TCGAG
PalI GG!CC
PflMI CCANNNN!NTGG
PleI GAGTCNNNN!
PleI !NNNNNGACTC
PmaCI CAC!GTG
PpuMI RG!GWCCY
PstI CTGCA!G
PvuI CGAT!CG
PvuII CAG!CTG
RsaI GT!AC
RsrI G!AATTC
RsrII CG!GWCCG
SacI GAGCT!C
SacII CCGC!GG
SalI G!TCGAC
Sau3AI !GATC
Sau96I G!GNCC
SauI CC!TNAGG
ScaI AGT!ACT
ScrFI CC!NGG
SduI GDGCH!C
SecI C!CNNGG
SexI CTCGAG
SfaNI GCATCNNNNN!
SfaNI !NNNNNNNNNGATGC
SfiI GGCCNNNN!NGGCC
SinI G!GWCC
SmaI CCC!GGG
SnaBI TAC!GTA
SnaI GTATAC
SpeI A!CTAGT
SphI GCATG!C
SplI C!GTACG
SspI AAT!ATT
SstI GAGCT!C
SstII CCGC!GG
SstIII ACGT
StuI AGG!CCT
StyI C!CWWGG
StySJI GAGNNNNNNGTRC
StySJI GYACNNNNNNCTC
TaqI T!CGA
TaqII GACCGANNNNNNNNNNN!
TaqII !NNNNNNNNNTCGGTC
TaqII CACCCANNNNNNNNNNN!
TaqII !NNNNNNNNNTGGGTG
ThaI CG!CG
Tsp45I GTSAC
TspEI AATT
Tth111I GACN!NNGTC
Tth111II CAARCANNNNNNNNNNN!
Tth111II !NNNNNNNNNTGYTTG
TthHB8I T!CGA
VspI AT!TAAT
XbaI T!CTAGA
XcyI C!CCGGG
XhoI C!TCGAG
XhoII R!GATCY
XmaI C!CCGGG
XmaIII C!GGCCG
XmnI GAANN!NNTTC
XorII CGAT!CG
26- نشان دار كردن اسيد هاي نوكلئيك
مقدمه: نشاندار كردن اسيد هاي نوكلئيك تحول بزرگي در بيولوژي مولكولي ايجاد نمود. اين تكنيك براي تاييد كارهاي مولكولي و همچنين غربالگري كلني هاي باكتري حاوي پلاسميد نوتركيب ( هيبريداسيون ) استفاده مشود. انشاا…. در اين كارگاه با روشهاي نشاندار كردن اسيد هاي نوكلئيك بصورت عملي آشنا خواهيم شد و روش هيبريديزاسيون را عملا" انجام ميدهيم. دراينجا مختصري از كارهايي كه انجام خواهد شد توضيح داده ميشود.
رو شهاي نشاندار كردن آنزيماتيك DNA :
Nick Translation
سنتز پروب بكمك .PCR
در nick translation نوكلئوئيدهاي نشاندار نشده با نوكلئوئيدهاي نشاندار شده تعويض ميشوند) سنتزجابجايي.( در دو رو ش ديگر منجربه سنتز DNA جديد مي شود (Net synthesis)
27- Random - primed labeling
در هيبريديزاسيون علاوه بر استفاده از پروب هاي نشاندار شده با راديوايزوتوپها ازگروههاي تغيير يافته غير راديواكتييو مانند بيوتين ياديگوكسي ژنين نيز استفاده ميشود .اين رو ش در سال 1984 و 1983توسط Feinberg و Vogelstein ابداع گرديد .نشانداركردن توسط آنزيم klenow انجام ميشود كه قطعه بزرگ DNA پلميراز I اشرشياكلي ميباشد. Klenow فاقد فعاليت اگزونوكلئازي 5` 3` ميباشد ولي برعكس داراي فعاليت اگزونوكلئازي 3` 5` ميباشد. براي نشاندار كردن با روش Random - Priming اول DNA دو رشتهاي )خطي( توسط حرارت دناتوره شده و سپس روي يخ سرد ميشود تامانع رناتوراسيون DNA شده ويصورت زنجيره ها ي تك رشته اي باقي بماند. Supercoiled DNA مقدار كمتري نشاندار ميشود چون Reannealing رشته هاسريعتر انجام ميشود.بنا براين مولكولهاي DNA حلقوي بايد قبل از نشاندار شدن بصورت خطي درآمده باشند تا عمل Labeling به مقدار زياد انجام گيرد.از قطعات كوچك DNA هگزا نوكلئوتيدي با تواليهايمتفاوت به عنوان پرايمر(آغازگر (براي سنتز DNAبكمك يك آنزيم DNA پليمراز استفادهميشود .مخلوط هگزانوكلئوتيدهابه روش تصادفي – اتفاقي به DNA دناتوره شده هدف Anneal ميشوند. اين اليگونوكلئوتيدهادر واكنش پليمرازبعدي به عنوانprimer عمل ميكنند. سنتز پروب با اضافه كردن آنزيم پليمراز klenow و چهاردزوكسي نوكلئوتيد تري فسفات(كه حداقل يكي ازآنها (dUTP) باهاپتن نشاندارشده است) شروع ميشود .چون پرايمرها خاصيت تصادفي - اتفاقي دارند ميتوانند به همه ترادف هاي هدف ا تصال يابند . در اثناي سنتزبه روش Random - primed هر دو زنجيره DNA )رشتههاي مكمل DNA هدف (بعد از دناتوراسيون به عنوان الگو عمل ميكنند .غلظت زيا'د پرايمر مانع Reanealing دورشته الگو با يكديگر ميشود.در اين روش به علت عمل پرايمرهاي كوتاه معمولا" رشته DNA نشاندار كوتاهتر از DNA الگو مي باشد .چون هر دو رشته DNA اصلي به عنوان Template عمل ميكنند رشته هاي پروب نشاندار نيز تا اندازهاي مكمل يكديگر هستند، بطوري كه قبل از هيبريديزاسيون لازم است دناتوره شوند.از روش Random - primedبراي نشاندار كردن قطعات DNA به مقدار چند نانوگرم تا چندين ميكروگرم استفاده ميشود .نشاندار شدن بعداز 30ـ60 دقيقه به حداكثر ميرسد و هنگام انكوباسيون طولاني ورود ماده نشاندا ر به رشته ثابت است . حساسيت ها’پتنهايي مانند بيوتين و Digoxigeninاز طريق Random priming تا 100فمتوگرم DNA مبباشد.
28- Nick translation
اين روش اولين با ربراي نشاندار كردن پروب با را ديوايزوتوپ توسط Rigny در سال 1977 ابداع گرديد. در اين روش عمل دو آنزيم DNase I و DNA polymerase I اشرشياكلي روي DNAدو رشتهاي همزمانانجام ميگيرد. DNase I يك اندونوكلئاز اختصاصي هيدروليز كننده باند هاي فسفودي استر رشته هاي DNA ميباشد. اين عمل منجربه ايجاد اليگونوكلئوتيدهاي كوتاه حاوي 5` فسفات ميشود. عمل آنزيم روي هر رشته DNA مستقيما و در حضور يون Mg2+انجام ميگيرد و شكافهايي در DNAيك رشتهاي بوچود مي آورد كه تعداد آنها به غلظت Dnase Iدر مخلوط انكوباسيو ن بستگي دارد.اين عمل با غلظت پايين Dnase I هم انجام ميشود . در اين شرايط فقط-تعداد كمي Nick در هر زنجير DNA به وجود ميآيد .آنزيم DNA Polymerase I داراي سه فعا'ليت كاتا'ليتيك مستقل يعني پليمرازي 5` 3®` , اگزونوكلئازي 5`3® ` واگزونوكلئازي 3`® 5` ميباشد..فعاليت نوكلئاز ي 5` 3®` آنزيم باعث حذف دزوكسي نوكلئوتيدها از انتهاي 5` فسفات ميشود، با فعاليت پليمرازي 5` 3® ` آن ، آنزيم مذكوربطور همزمان نوكلئوتيدتري فسفاتها را به انتهاي 3`-OH آ زاد شكاف (Nick) ا'ضافه ميكند .اگر دزكسي نوكلئوتيد تري فسفات هاي همراه دزوكسي نوكلئوتيدتري فسفاتهايتغيير يافته با ها'پتن در مخلوط واكنش با شند در اثناي واكنش پليمريزاسيون هاپتن وارد زنجيره ميشود .چون عمل DNA Polymerase I وابسته به DNA الگو است ترادف نوكلئوتيدي هنگام Nick translation بدون تغييربا قي ميماند.
29- نشاندار كردن DNA توسطPCR
PCRيك روش توانمند براي سنتز پروبهاي DNA ( DNA بدون وكتور (مي'باشد اين روش در سال1984توسط موليس ابداع گرديد وبراي كارهايي مانند تكثير, , Cloning Sequencingو سنتز پروب بكار گرفته شد .در اين روش دو پرايمرحاوي دزوكسي نوكلئوتيدتري فسفاتهاي نشاندار شده درپليمريزاسيون DNA شركت ميكنند PCR .توسط يك DNA پليمراز گرمادوست-مانند Taq انجام ميشود .واكنش Amplification در30 سيكل حرارتي در سه مرحله دناتوراسيون، Annealingو Extenssion انجام ميگيرد.
30- هيبريديزاسيونHybridization
DNAدو رشتهاي محلول تحت تاثير حرار ت دناتوره ميشود .اگر حرار ت در حدTm مولكول DNA باشد دناتوراسيون برگشت ناپذير است يعني رشتههايDNA در محيط سرما جدا از يكديگر باقي ميمانند Tm .يك مولكول DNA دو رشتهاي ايستايي DNA در حرارت مي'باشد .(Function of the Thermal Stability) در يك DNA هومولوگ اين پديده انعكاس نسبت با زها يعني محتواي G+C موجود در DNA ميباشد. Cو Gبا سه اتصال هيدروژني بهم متصل ميشوند در حاليكه Tو A با دو اتصال بهم مربوط ميشوند Tm .مولكولDNA تحت تاثير قدرت يوني محيط (با افزايش يونهاTm بالا ميرود(، حضور يونها(Mg+) ، پلي ساكاريدها, پرتئين و حلالهاي آلي (Formamide) قرار ميگيرد .رشتههاي DNA دو رشتهاي دناتوره شده در حرار ت كمتر از Tm خودبخودبهم چسبيده و نهايتابا DNA مكمل خود يك هيبريد پايدارتشكيل ميدهند بنا بر اين واكنش از كينتيك ثانوي (Second- Order Kinetics) تبعيت ميكند. هيبريديزاسيون نيز شبيه Tm تحت تاثير فاكتورهايي مانند قدرت يوني و حلالهاي آلي قرار ميگيرد. اصطلاح Stringency در اين مورد مفهوم مهمي است اما همه اين فا'كتورها )حرارت ، غلظت نمك، وجود حلالهاي آلي( را دربرنميگيرد .در شرايط Stringency پايين )نمك زياد و حرارت كمDNA ( هاي با شباهت كمتر با همديگر هيبريد ميشوند .در صورتيكه درStringency بالا) غلظت كم نمك و حرارت بالا در حضور فرماميد( Hybridization فقط بين دو DNA باهومولوژي با لا اتفاق ميافتد .در اين شرايط حرارت Hybridization30ـ20 درجه پايين تر از (melting temprature) tm است .براي انجام Hybridizationبايد DNA هدف را روي يك پشتيان جامد منتقل نمود تا DNA نشاندار (بصورت محلول) با DNA هدف هيبريد تشكيل دهند .در ساترن بلات DNA يا RNA روي ژل آگارز الكتروفورز ميشوند و سپس روي غشاي نيتروسلولزيا نايلوني منتقل ميگردند .از مواد پشتيان كننده ديگر مانند سلولز فعال، سفاكريل، پلي استيرن يا بيدهاي مگنيتك نيز ميتوان استفاده نمود.
پارامترهايHybridization
پايداري هيبريد:
تشكيل هيبريدهاي اسيدنوكلئيك يك پروسه برگشت پذير است . Tm عبارت است از حرارتي كه نصف مولكولهاي DNA دوپلكسبه تك رشته تبديل ميشوند و تحت تاثيرغلظت نمك كاتيونهاي يك ظرفيتي) مول بر ليتر (تعداد نوكلئوتيدهاي زنجيره، تركيب'بازها ( كه با G+C بيان ميشود (و غلظت عوامل ناپايدار كننده هليكس مانند Formamideقرار ميگيرد.
كينتيك هيبريديزاسيون پروبهاي تك رشتهاي: (Riboprobe)
ميزان تشكيل هيبريد براي پروبهاي تك رشتهاي از كينتيك اوليه (First - Order Kinetics)تبعيت ميكند زيرا تقريبا هميشه غلظت پروب بيشتر از Target DNAاست ، بالاترين ميزان هيبريديزاسيون در محلول بصورت تجربي مشخص ميشود. درجه Tm در دو پلكس DNA- DNA 15 درجه كمتر از دوپلكسRNA-DNA است. غلظت كاتيون (M) تاثير كمتري روي ميزان (Constant rate) ثابت هيبريديزاسيون DNA-DNAدارد .ميزان هيبريديزاسيون تحت ثير قدر ت يوني پايين قرار ميگيرد. هيبريديزاسيون در 1M NaCl هفت برابربيشتر از 0.18M NaCl انجام ميگيرد .كاهش غلظت نمك در حد 0.09M NaCl تا 5 برابر ميزان هيبريديزاسيون را كاهش ميدهد. تا ثير نمك روي تشكيل دوپلكس RNA- DNA تا اندازهاي با آنچه ذكر شد اختلاف دارد .در 0.1M NaC تشكيل DNA-RNA همسنگ DNA-DNA (Equivalent) است ولي در 1M NaCl ميزان تشكيل دوبلكس RNA-DNA دوبرابر 0.18M است.
كينيك هيبريديزاسيون در پروبهاي دو رشتهاي (DNA)
در پروبهاي دو رشتهاي هيبريديزاسيون از كينتيك ثانويه (Second - Order Kinetics)تبعيت ميكند. اين پروبهامي توانند بااسيدهاي نوكلئيك ثابت شده روي فيلتر هيبريد شوند و يادر محلول دوباره Renature گردند. در نتيجه هيبريديزاسيون 5 برابر آهسته تر از پروبهاي تك رشتهاي انجام ميگيرد. پليمرهاي دكستران آنيوني) دكستران سولفات 500) ياپلي اتيلن گليكول 6000 اتصال دو رشته پرو ب را در محلول به همديگرتشديد ميكنند .دكستران سولفات محلول را از DNA خارج ميكند بنا براين غلظت موثر DNA افزايش مييابد .اثر دكستران سولفات براي پلي نوكلئوئيدهاي بيشتر از250bp مشهودتر است و روي اليگونوكلئوئيدها تاثيري ندارد. وقتي از پروبهاي تك رشته اي استفاده ميشودهيبريديزاسيون تا سه برابر افزايش ميابدو وقتي از Nick translated probe استفاده شود تا 100برابر ميشود .مولكولهايي كه از طريق Nick translation نشاندار شدهاند دكستران سولفات تشكيل شبكه پروب (probe network)يا hyper polymer هاراتسريع نميكند وتاكيد ميشود كه تشكيل چنين شبكههايي به اندازه پرو ب بستگي دارد. خاطر نشان ميسازد كه اگر دكستران سولفات در محيط نباشدbackground هاي غيراختصاصي زياد ميشوند.
روش كار:
براي نشاندار كردن قطعه DNA ( پروب) از كيتRandom Primed DNA Labeling شركت Roch Molecular Biochemicals كه با سيستم Digoxigenin (غير راديواكتيو) كار ميكند استفاده ميشود. ديگوكسي ژنين يك هاپتن استروئيدي است كهDNA ، RNAو اليگونوكلئوئيدها را جهتHybridization نشاندار ميكند. و ظهور(Detection) آن بصورت كالريمتري انجام ميشود. براي نشاندار كردنDNA ، ديگوكسي ژنين از طريق اتصال استري وارد dUTP ميشود، اتصال فوق دربرابر قليا (alkali) ناپايداربوده و اين يك امتيازي است كه DIG - 11 - dUTP به آساني توسط قليااز رشته مكمل روي Filter ( كاغذ نيتروسلولز يا غشا’ نايلون) شسته شده و ميتوان دوباره از فيلتربراي هيبريديزاسيون با پرو ب ديگر استفاده نمود.
ـ DNA را مد ت 10 دقيقه در آ بجو ش قرار داده دهيد تا به Single strand DNA تبديل شودوبلافاصلهان را روي يخ حاوي نمك منتقل كنيد و مواد زيررا به آن اضافه نماييد.
ـ 2 ميكروليتر ازمخلوط هگزانوكلئوئيدها
- 2 ميكروليتر مخلوط.dNTP
- آب مقطر تا حجم 19 ميكروليتر
- 1 ميكروليتر آنزيم klenow
با دست به ته لوله ضربه بزنيدتا مواد داخل لوله مخلوط شوندو مختصري سانتريفوژكنيد (quick spin)
- مد ت 24 ساعت (O/N) در 37 درجه قراردهيد.
- واكنش رابا EDTA متوقف كرده وبراي پرسيپيتاسيون Licl با غلظت نهائي 0.5 M به آن اضافه كنيد
-رسوب حاصل را در 50 ميكروليتر بافر TE حل كنيد.
ـ براي كنترل Labeling يك واكنش نيزبا DNA كنترل موجود در كيت انجام دهيد.
بررسي كمي و كيفي پروب نشاندار شده:
از پروب نشاندار شده رقت سريال( 10/1, 100/1, 1000/1) تهيه كنيد ( به هر يك از ويالها 9 ميكروليتر آب اضافه كرده سپس يك ميكروليتر از پروب به لوله اول اضافه كرده و خوب مخلوط كنيد و همينطور يك ميكروليتر از لوله اول به لوله دوم منتقل كنيدو……..) با انجام Dot blotروي نايلون يا نيتروسلولز آنهارا ظاهر (detect) كنيد.
- پس از انجام (dot) لكهگذاري قبل از اينكه لكهها'كاملا خشك شوند توسطUV Crosslinker ( مقدار 12000 ژول انرژي) آنها را روي غشا ثابت كنيد ( وقتي DNA در معرض UV قرار گيرد تيمين موجود در آن با گروههاي آمين روي سطح نايلون cross-link ميشوند .
- نايلون را مدت 10 دقيقه دربا فر I قرار دهيد. غشا را مدت 1 ساعت دربا فر II قرار دهيد تا فيلتر block شود( جاهائيكه DNA وجود ندارد توسط BSA پوشيده (block) ميشود تا آنتي بادي نتواند روي غشا بچسبد)
- غشا را مدت 20 دقيقه در Anti Dig بارقت 5000/1 قرار دهيد.
- غشا را چند دفعه با بافر I شستشو داده تا آنتي با ديهاي متصل نشده از روي فيلتر حذف شوند.
- غشا را بابافر III شستشو دهيد
- لكههاي روي غشا را با (Nitro Blue Tetrazolium (NBT و (Bromo Chloro Indolyl Phosphate (BCIP ظاهر كنيد
ـ با مقايسه رنگ لكهها بارنگ لكه DNA كنترل نشاندار شده موجود در كيت مقدار پروب نشاندار شده را محاسبه كنيد). هر ميكروگرم DNA كنترل حاوي ... نانوگرم DNA نشاندار شده ميباشد.
تهيه Riboprobe
ـ داراي طول معيني بوده و اختصاصي وتك رشتهاي مي با شند.
شبيه DNA probe ها به يكديگر متصل نميشوند.
ـ اتصال Dig به UTP نسبت به NaOH مقاوم ميباشد.
رو ش كار:
ـ بعد از اتمام واكنش آن را P.C.I extroction كرده سپس با الكل رسوب دهيد.(هرگزازRNase استفاده نكنيد).
-2 ميكروليتر NTP به واكنش اضافه كنيد.
- 2 ميكروليتر 10X Transcription Buffer به آنا ضافه كنيد.
ـ 1 ميكروليتر ((T3, T7) RNA ploymerase به آن اضافه كنيد.
- حجم واكنش را با آب مقطربه 20 ميكروليتر برسانيد.
-1 ميكروليتر RNasin ( مهار كننده RNase ) به واكنش اضافه كنيد تااز فعاليت RNase احتمالي درواكنش جلوگيري كند.
- لوله را مختصري سانتريفيوژ (quick spin) نموده و 2ـ1 ساعت در 37 درجه قرار دهيد.
- واكنش رابا EDTA متوقف نموده وبراي رسوب دادن RNA ازLiCl باغلظت نهائي 0.5M استفاده كنيد.
- بعد از شستشو با الكل 70، رسوب را در 100 ميكروليترDEPC treated water حل كنيد.
- مقدار 1 ميكروليتر RNasin به آن اضافه كرده و 30 دقيقه در 37 درجه انكوبه كنيد.
- كميت پروب توليد شده شبيه DNA probe تعيين ميشود فقط براي تهيه بافر II ازDEPC treated water استفاده شود
- از RNA نشاندار شده موجود در كيت براي مقايسه پروبها استفاده كنيد.
Hybridization
روش دات بلات
- مدت 5 دقيقه غشانايلوني را در محلول Denaturing قرار دهيد تا DNA دناتوره شود.
( لازم به ذكر است كه نايلون نبايد در محلول شناور شود بلكه بايد ته ظرف يا سيني يك لايه كاغذ خشك كن قرار داده شود سپس محلول را روي كاغذ ريخته بطوري كه فقط كاغذ خيس شود وسپس نايلون را روي آن قرار دهيد.
ـ 5 دقيقه غشارا مانند مرحله قبل در محلول Neutralizing قرار دهيد تا.غشا خنثي شده و از شكنندگي آن جلوگيري شود.
- غشا را روي كاغذ خشك كن قرار داده و هنگا ميكه هنوز مرطوب است DNA را باUV Cross Linker روي أن ثابت كنيد.
ـ غشا در اين مرحله آماده است هم ميتوان براي مدتي آن را نگهداري نمود و هم ميتوان پروسه Hybridization را دنبال نمود.
روش ساترن بلات
ـ وقتي الكتروفورز انجام شد ژل رابا UV مشاهده كرده وبراي جلوگيري از اصراف مواد قسمتهائي از ژل را كه مورد استفاده نيست حذف كند( ژل را Trim كنيد) .
- گوشه( يكي از گوشهها ) ژل را علامت گذاري كرده و سپس 90 دقيقه در حرارت اتاق آن را داخل محلول Denaturing دوران دهيد تا DNA دناتوره شود (0.4N NaOH) البته DNA هاي سنگين تراز15kb را بايد با محلول0.25 M اسيد كلريدريك Depurinate نمود تا بتوانند از ژل روي غشا منتقل شوند (اين كار همراه باshaking انجام شود).
-واكنش را 20 دقيقه در حرارت اتاق در محلول Neutratizing قرار دهيد ( همراه باshakingانجام گيرد).
- غشا (Nylon membrane) رابه اندازه ژل بريده و گوشه آن را مانند ژل علامت گذاري كرده و سپس در آب ديونيزه آن را خيس كنيد.( 98).
- بعد از آماده كردن تا نك انتقال (Transfer Tank) يك قطعه كاغذ صافي روي سيني تا نك قرار دهيد.
- كاغذ صافي را حتما" بابافرتا نك خيس كنيد كه حباب هوا در آن قرار نگيرد .
- يك لايه كاغذ واتمن3MM روي آن قرار دهيد بطوري كه دو سر كاغذ دربا فرتا نك قرار گيرد.
- ژل رابصورت وارونه( نسبت به موقعي كه الكتروفورز ميشود ) روي كاغذ واتمن قرار داده بطوري كه زير ژل حباب هوا قرار نگيرد.
- نايلون آماده شده را روي ژل قرار دهيد بطوريكه هوا زير آن نفوذ نكند.
ـ يك لايه كاغذ واتمن روي فيلتر قرار دهيد.
- اطراف آن را با پارافيلم خوب بپوشانيد.
ـ يك لايه كاغذ صافي و سپس چندين لايه كاغذ خشك كن روي آن قرار دهيد بطوري كه مايع را جذب كند.
- يك صفحه شيشه اي روي آن قرار دهيدويك وزنه 500 گرمي روي شيشه مستقركنيد.
- اطراف تا نك را با نايلون بپوشانيد كه تبا دل هوا با خارج انجام نگيرد واز تبخيربافرتا نك ممانعت نمايد.
- براي بافرتا نك از محلول 0.4N NaOH استفاده كنيد زيرا براي Nylon بهتر از كاغذ نيتروسلولز ميباشدو موجب دناتوره شدن و ثابت شدن DNA روي نايلون ميشود.
- مدت 24 ـ4 ساعت در هواي اتاق قرار دهيد.
- نايلون را خارج كرده و با 2X SSC بشوئيد تا ذرات ژل ازروي آن حذف شوند.
- وقتي نايلون هنوز نمدار است DNA را با دستگاه UV crosslinker روي آن ثابت كنيد.
Pre hybridization ( دات بلات يا'ساترن بلات )
ـ غشارا داخل كيسه پلاستيكي (Hybridization bag) قرارداده ومقدار 20ml/100cm2 محلول Prehybridization داخل كيسه ريخته و سپس آن را خوب درز گيري كنيد بطوريكه مايع از آن نفوذ نكند( seal شود).
ـ مدت 2- 5/1 ساعت در 42 درجه قراردهيد( براي ريبوپروب حرارت 50 درجه درنظرگرفته شود )
ـ پروب را مدت 5 دقيقه درآب جوش قرار داده تا دناتوره شود
- بلافاصله روي يخ حاوي نمك قرار دهيد.
ـ يكي از گوشه هاي كيسه پلاستيكي حاوي nylon را بريده و آن را روي كاغذ خشككن قرار داده و با چرخانيدن يك مداد يا ميله شيشهاي گرد روي آن , محلول Prehyridization را از آن خارج كنيد.
- مقدار 2.5ml/100cm2 پروب مخلوط شده با محلول Hybridization وارد كيسه كنيد.
- هواي آن را خارج كرده دوباره آن را seal كنيد.
- مدت يك شب آن را در 42 درجه قرار دهيد. ( براي ريبوپروب 50.درجه).
Post hybridization
- مدت 5 دقيقه بامحلول2XSSC , 1% SDS همراه shaking در حرارت اطاق نايلون را بشوئيد.
- مدت 15 دقيقه بامحلول 2XSSC , 1% SDS آن را يشوئيد (RT)
- مدت 30 دقيقه بامحلول 2XSSC , 1% SDS در65 درجه شستشو دهيد.
- Detection آن شبيه پروب انجام ميگيرد.
هيبريديزاسيون با Riboprobe
- مراحل Dot blot يا Southern blot شبيه DNA پروب انجام ميگيرد.
ـ دماي Hybridization را 50 درجه درنظربگيريد.
– بهتر است پروب (RNA) را هنگام استفاده مدت 5 دقيقه در آب جوش قرار دهيد تا چنانچه RNAدايره (Loop) تشكيل داده است باز شود.
ـ مراحل شستشو و Detection نيز شبيه DNA انجام ميگيرد.
ـ بايد دقت نمود كه حين پروسه كار Rnase وارد محيط نشود وازتما'س دست يا با وسايل كار و نمونه خودداري شود.
- حتما" از دستكش استفاده كنيد
- وسايل شيشهاي، تيپها و محلولهارا با يد از آلوده شدن با RNase محافظت نمود.
- موادووسايل كاربا RNA بايد از وسايل و مواد ديگر جداباشند. وسايل شيشهاي بعد از شستشوبا يد در محلول 1/0درصدDEPC شناور شوند و سپس اتوكلاو گردند وبعد از آنبمدت 3 ساعت در250 درجه قرار گيرند.
تيپها ، لولههاي سانتريفيوژ و لولههاي فالكن فاقد RNase ميباشند ولي با يداتوكلاو شوند.
DEPCماده مهار كننده غير اختصاصي ريبونوكلئاز ميباشد كه با آدنين موجود درRNA واكنش ميدهد.
تهيهDeionized formamide
روش كار:
- كاتيون و آنيون( از هر كدام 5 گرم) جداگا نه بايك ليترآب Equilibrate ميشوند. بعد از مخلوط شدن محلول ابري مانند تشكيل ميگردد .اين محلول را با دستگاه فيلتراسيون در خلا فيلتر( كاغذ صافي واتمن ) كرده تا آب رزين خارج شده و رزين روي فيلتربا قي بماند و خشك شود سپس با يك ليتر فرماميد 30 دقيقه روي همزن ميچرخد تا خوب مخلوط شود .محلول دو باره با دستگاه فيلتراسيون درخلا با كاغذ صافي واتمن فيلتر ميشود بطوري كه يك محلول صاف تشكيل ميگردد.اين محلول Deionized Formamideدر يخچال نگهداري ميشود.
طرزتهيه بافرها:
1- بافرI:
Tris (PH:7.5) 100mM
NaCl 500mM
2- بافرII :
(1-3% BSA (blocking reagent كه در بافر I 65درجه حرارت داه ميشود تا حل شود
3- بافر III:
Tris (pH:9.5) 100mM
NaCl 100mM
MgCl2 50mM
ب) بافرهاي Hybridization :
1- بافر Pre hybridization:
Deionized formamide 50%
SSC 5X
Denhardts 5X
hosphate buffer 50mM
SS DNA 125mg/ml
2- بافر Hybridization :
Deionized formamide 50%
SSC 5X
Denhardts 1X
Phosphat buffer 20mM
SS DNA 25mg/ml
Dextran sulfate 10%
3- محلول Denhardts :
BSA 1%
Ficoll 1%
Polyvinyl pyrolidone 1%
4- با فر 20X SSC :
NaCl 3M
Na Citrate 3M
pH: 7
منبع:http://www.iranbiotech.com
انجام واكنش Ligation:
2- حجم واكنش را در نظر بگيريد ( بهتر است يك ميكروگرم DNA را درواكنشي به حجم 20-30 ميكروليتر انجام گيرد )
3- با آب دوبار تقطير استريل حجم واكنش را تنظيم كنيد
4- واكنش را مدت 5 دقيقه در 45 درجه قرار دهيدتا انتهاهاي Cohesive كاملا" Relax شوند
5- بافر Ligation را با غلظت نهايي 1x استفاده كنيد
6- يك ميكروليتر آنزيم T4 DNA ligase به واكنش اضافه كنيد
7- چند ثانيه آن را spin كنيد
8- مدت 3-2 ساعت در دماي 37 درجه قرار دهيدو سپس واكنش را ترانسفرم كنيد.
18- پيدا كردن كلني هاي حاوي Recombinant DNA:
در اين كارگاه آموزشي از دو روش استفاده ميكنيم
الف ) غير فعال كردن ژن مقاومت به تتراسيكلين پلاسميد pBR322
جايگاه آنزيم BamHI روي سكانس ژن مقاومت به تتراسيكلين پلاسميد pBR322 قرار دارد چنانچه DNA خارجي در جايگاه اين آنزيم كلون شود و پلاسميد نو تركيب در باكتري ترانسفرم گردد ، باكتري نسبت به تتراسيكلين حساس ميشود.Insert DNA مورد استفاده كه قطعه اي از DNA ميتوكندري (kDNA) انگل ليشمانيا ميباشد در حدود 800bp است كه آن را در جايگاه BamHI پلاسميد pBR322 كلون ميكنيم
1- محصول Ligation را درباكتري HB101 ترانسفرم كنيد ( باكتري XL1-blue نسبت به تتراسيكلين مقاوم است و براي اين آزمايش قابل استفاده نميباشد).
2- محصول ترانسفرم شده را روي پليت آگار حاوي آمپي سيلين و بدون تتراسيكلين پخش كنيد.
3- روز بعد يك Master plate حاوي آمپي سيلين و بدون تتراسيكلين تهيه كنيد و آن را شطرنجي نموده و خانه هاي آن را شماره گذاري نماييد سپس كلني هاي حاصل از ترانسفرماسيون شب قبل را داخل خانه هاي شطرنجي (شماره دار) پليت منتقل كنيد.
4- روز بعدنيز آگار پليت مشابه روز قبل تهيه كنيد كه داراي دو آنتي بيوتيك آمپلي سيلين و تتراسيكلين باشد (100μg /ml آمپي سيلين و 12.5μg/ml تتراسيكلين ) و هر كلني را در خانه هم شماره آن روي پليت حاوي تتراسيكلين و آمپي سيلين كشت دهيد
5- روز بعد هر كدام از كلني ها كه رشد نكرد ه باشند حاكي از حساس بود ن آنها به تتراسيكلين ميباشد (قطعه DNA مورد نظردر پلاسميد كلون شده است).
6- كلني باكتري كه رشد نكرده را از روي Master plate شماره 1 ( بدون تتراسيكلين ) پيدا كرده و از آن كشت شبانه تهيه كنيد
پلاسميد آن را استخراج كرده و با آنزيم BamHI آن را برش دهيد ، بعد از الكتروفورز بايد قطعه DNA كلون شده را روي ژل مشاهده كنيد ( تقريبا" 800bp ميباشد ) .
ب) غير فعال كردن ژن LacZ` پلاسميد Bluescript و مشاهده كلني ها ي آبي و سفيد
( alfa- complementation test ) همانطور كه قبلا" گفته شد MCS پلاسميد Bluescript داخل ژن LacZ` تعبيه شده است . سكانس اين ژن قسمتي ازسكانس ژن آنزيم β-galactosidase است , اين آنزيم اثصالات بتا گالاكتوزيدي لاكتوز را تجزيه كرده و لاكتوز را به گلوكز و گالاكتوز تبديل ميكند , همچنين ماده X-gal را كه آنالوگ لاكتوز ميباشد تجزيه كرده و رنگ آبي توليد ميكند. سلولي (باكتري) كه آنزيم بتا گالاكتوزيداز آن ناقص باشد يعني ژن قطعه LacZ` را نداشته باشد چنانچه با پلاسميدي كه سكانس ژن LacZ` داشته باشد ترانسفرم گردد آنزيم بتا گالاكتوزيداز آن كامل ميشود و ميتواند لاكتوز يا X-galرا تجزيه كند. چنانچهinsert DNA درMCS اين پلاسميد كلون شود ژن مذكور غير فعال شده و باكتري نميتواند بعد ترانسفرم شدن با پلاسميد X-gal را تجزيه كند در نتيجه رنگ آبي توليد نميشود و كلني هاي بيرنگ توليد ميشود
A) در سيستم alfa-complementation test. كروموزوم باكتري ژن lacZ ناقص دارد و قسمت N ترمينال(alfa peptide) ژنβ-galactosidase راكد نميكند و محصول ژن فاقداين قسمت ميباشد . ژن lacZ` پلاسميد در داخل باكتري قسمت alfa peptide را كد ميكند و ژن بتا گالاكنوزيداز فانكشنال توليد ميشود كه در حضور X- gal رنگ كلني هاي باكتري آبي ميشود . (b يك قطعه DNA در پلاسميد كلون شده ,ژن lacZ` ناقص شده است ونميتواندX-gal راتجزيه كند در نتيجه كلني هاي باكتري سفيد ميگردند.
1- DNA را در جايگاه آنزيم BamHI يا EcoRI پلاسميد Bluescipt كلون ( Ligate ) كنيد.
2- واكنش Ligation را داخل باكتري XL1- blue ترانسفرم كنيد ( ژنآنزيم β-galactosidase اين باكتري ناقص است و همراه ژن LacZ` پلاسميد Bluescript كامل ميشود اما ژن آنزيم بتا گالاكتوزيداز باكتري HB101 كامل است وبراي اين منظور قابل استفاده نميباشد )
3- باكتري ترانسفرم شده را روي آگار پليت حاوي X-gal و IPTG پخش كنيد ( IPTG يك ماده القاء كننده پروموتور ژن است و موجب بيان آنزيم β-galactosidase ميشود ) .
4- قبل از پخش كردن واكنش روي پليت , به هركدام مقدار 40 ميكروليتر از 20 mM X- gal و 4 ميكروليتر از 200 mg/ml IPTG اضافه كنيد.
5- روز بعد ( 16-12 ساعت ) كلني هاي آبي و سفيد روي پليت آگاررشد ميكنند كه كلني هاي سفيد حاوي Recombinsnt plasmid ميباشند
توجه : اگر پليت حاوي X- gal و IPTG را چند ساعت در 4 درجه قرار دهيد رنگ آبي بخوبي ظاهر ميشود. مركز كلني حاوي ژن بتا گالاكتوزيداز آبي كم رنگ و محيط آن آبي پررنگ است. در مركز كلني هاي سفيد نقطه آبي خيلي كم رنگ ديده ميشود ولي محيط آنها بيرنگ است.كلني هاي سفيد راكشت دهيد و پلاسميد آنهارااستخراج كنيد. پلاسميد را با آنزيم BamHI هضم كنيد بايد بعد از الكتروفورز قطعه DNA كلون شده را روي ژل مشاهده كنيد
19- Polymerase Chain Reaction ( PCR)
واكنش زنجيره اي پليمراز روشي است كه قسمتي از سكانس مولكول DNA كه بين دو پرايمر قرار دارد توسط آنزيم پليمراز وبكمك چهار داكسي نوكلئوتيد تري فسفات در آزمايشگاه تكثير (Amplify) ميشود. .dsDNA متشكل از دو رشته آنتي پارالل ميباشد كه توسط اتصالات هيدروژني وبصورت كووالانت به يكديگر متصل هستند. همانند سازي DNA بكمك اليگو نوكلئو تيد هايي كه پرايمر گفته ميشوند انجام ميگيرد. اليگونوكلئوتيدها مولكولهاي DNA تك رشته كوتاه هستند كه هر كدام از آنها مكمل يك انتهاي سكانس DNA هد ف ( الگو ) مي باشند.پرايمر ها توسط آنزيم DNA پليمراز و در حضور dNTPها از روي DNA الگو ( تك رشته اي است ) همانند سازي ميكنند و رشته هاي جديدي مكمل رشته هاي هدف سنتز ميگردد.
مراحل يك سيكل PCR :
1- مرحله Denaturation : در اين مرحله DNA هدف بر اثر حرارت تك رشته اي ميشود.
2- مرحله Annealing : در اين مرحله با كاهش حرارت سيستم ، پرايمر ها در محل مناسب روي رشته مكمل متصل ميشوند
3- مرحله Extension : در اين مرحله كه دماي آن براي آنزيم DNA پليمرازمطلوب ميباشد موجب توسعه پرايمر ها شده و همانند سازي DNA هدف انجام ميگيرد. محصول PCR عبارت است از قطعه همانند سازي شده اي كه دو طرف اين قطعه سكانس پرايمر ها وجود دارند .
فاكتور هايي كه روي كار آيي PCR تاثير دارند:
1- بعد از 25تا 30 سيكل بعلت حرارت آنزيم دناتوره شده و غير فعال ميشود
2- غلظت زيادرشته هاي هدف موجب Reannealingآنها شده و با پرايمر ها رقابت ميكنند .
روش PCR توسط موليس كارمند شركت Cetus ابداع گرديد. ابتدا اين كار توسط سه بن ماري با حرارت هاي مختلف انجام ميگرفت واز آنريم كلنو بعنوان DNA polymerase استفاده ميشد. اين آنزيم در اثرحرارت دناتوره ميشودو اجبارا"بايد دوباره در هر سيكل به واكنش اضافه شود. Saiki از آنزيم DNA polymerase مقاوم به حرارت كه از باكتري ترموس آكواتيكوس خالص ميشود و اصطلاحا" Taq DNA polymerase گفته ميشود استفاده كرد وامروزه واكنشPCR بصورت اتوماتيك انجام ميگيرد.
پارامتر هاي موثر در PCR :
2- دماي Annealing پرايمرها كه بايد 3-4 درجه كمتر از دماي ذوب پرايمر ها باشد
3- زمانPrimer extension كه به تعداد بازهاي بين دوپرايمر بستگي دارد
4- - تعداد سيكلها
5- Ramp ( زماني كه طول ميكشد تا دماي مبدا دستگاه به دماي مقصد برسد ) هرچه اين زمان كمتر باشد نتيچه كار بهتر است و واكنش زمان كمتري در دماي ناخواسته قرار ميگيرد
6- غلظت dNTP ها و يون منيزيوم ( اينها مجموعه اي را تشكيل ميدهند كه موجب فعاليت آنزيم پليمراز ميشوند و غلظت اين دو ماده تابعي از يكديگر ميباشند )
7- غلظت Template DNA ( يك دهم تا يك ميكروگرم ميباشد ) چنانچه DNA هدف بصورت مالتي كپي در ژنوم باشد بهتر است.
8- اضافه كردن تشديد كننده هاي PCR
10- حذف مها ركننده هاي آنزيم از محيط
11- بهتر نقطه ذوب پرايمر ها (شبيه هم باشد ( Tm يكسان داشته باشند).
20- طراحي پرايمر :
پرايمر ها ي PCR بصورت كاملا" اختصاصي ومكمل ناحيه مورد نظر DNA هدف طراحي ميگردند. پرايمر ها 30-20 بازدارند و پرايمر هاي بيش از 30 باز اختصاصيت خوبي ندارند. همچنين پرايمر بايد دماي آنيلينگ بالا داشته باشد. بهتر است تعداد بازهاي دوپرايمر مساوي باشند و از پلي پورين يا پلي پيريميدين نباشند، همچنين نواحي تكرار شونده نداشته با شند چنانچه بازهاي G يا C بصورت تكراري و پشت سرهم باشند پرايمر بصورت لوپ در مي آيد و عملا" سيستم كار نميكند. انتهاي 3` دو پرايمر نبايد مكمل باشد زيرا پرايمر دايمر تشكيل ميشود.نرم افزارهايي وجود دارد كه طراحي پرايمر را انجام ميدهند. بعد از طراحي پرايمر بهتر است تو سط نرم افزارهايي مانند Blast آنها را چك نمود تا مشخص شود كه با چه ژنهاي ديكر ميتوانندAnneal گردند.در هر سيكل PCR تعداد مولكولهاي DNA دو برابر ميشود. رشته هاي DNA توسط حرارت (95-93درجه ) از يكديگر باز ميشوند ( Denaturing ) و سپس دردوره بعدكه حرارت پايين مي آيد (60-50درجه ) پرايمر ها بصورت اختصاصي به ناحيه هدف متصل ميشوند ( Annealing ). آنزيم Taq DNA polymerase در دماي مطلوب (72درجه ) ودر بافر مخصوص , چهار( dATP, dTTP, dCTP, dGTP ) dNTP را طبق رشته الگو بهم متصل ميكند (Extension ) .لازم به ذكر است كه DNA هاي كوتاه ( Short target product ) يعني رشته هايي كه بين دوپرايمر محصور هستند بصورت تصاعد هندسي (exponential ) و DNA هاي بلند ( long target product ) يعني رشته هايي كه از يك طرف توسط يك پرايمر محدود هستند و از سمت ديگر نامحدود ميباشند بصورت تصاعد حسابي (linearly ) زياد ميشوند .
مواقعي كه پرايمر ها كاملا" مكمل DNA هدف( الگو) نباشند :
1- پرايمر هايي كه براي ايجاد موتاسيون در يك ژن طراحي ميگردند
2- پرايمرهايي كه سمت 5` آنها جايگاه شناسايي آنزيم محدودگر تعبيه ميشود تا بتوان محصول PCR را توسط آن آنزيم برش داد . اين كار براي كلون كردن محصول PCR انجام ميشود.
3- پرايمر هايي كه لازم است محصول PCR آنها داراي پروموتور باشد و براي بيان يك ژن كاربرد دارند
فاصله دو پرايمر بايد كمتر از 10 kb باشد ( كارآيي همانند سازي براي محصول PCR بيش از 3kb كمتر است ) بهتر است پرايمر ها را بر حسب كاري كه لازم داريم طراحي شوند
چگونگي رد يابي ( detect ) محصول PCR :
1- هيبريد شدن محصول PCR با اليگو نوكلئوتيد نشاندار ( پروب )
3- الكتروفورز روي ژل آگارز يا پلي اكريلاميد
انواع PCR ( بمنظور تصحيح محصول ):
1- Hot start PCR : عبارت است از جدا كردن يك يا چند تركيب مهم PCR ( ترجيحا" انزيم پليمراز) بطوريكه بلا فاصله بعد ار دناتوراسيون DNA هدف به واكنش اضافه ميشوند ( استفاده از , PCR gem و آنتي بادي آنزيم پليمراز )
- Hot start بكمك آنتي بادي : مونوكلونال آنتي بادي ضد آنزيم پليمراز را به واكنش اضافه ميكننددرنتيجه فعاليت پليمرازي آنزيم را مهار ميكند. هنگاميكه دماي واكنش به 94 درجه ميرسد آتي بادي دناتوره ميشود و دو باره پليمراز فعال ميشود.
- Hot start بكمك Ampliwax : به ديواره لوله هاي محصوص اينكاربه نوعي واكس آغشته است كه بعد از مختصري گرم كردن ذوب شده و روي ئاكنش را ميپوشاند . آنزيم پليمراز را روي واكس قرار ميدهند . در دماي 94 درجه اين واكس بخار ميشود و آنزيم پليمراز با واكنش تماس پيدا ميكند
2- Touch down PCR : در اين روش دماي آنيلينگ ازدماي بالاتر از Tm بندرت كاهش ميابد وموجب كاهش محصول كاذب و پرايمر دايمر ميشود.
3- Nested PCR : مواقعي كه DNA هدف در نمونه مورد آزمايش كم باشد براي جلوگيري از بالابردن مقدار DNA و مهار واكنش توسط پرايمر هاي خارجي قطعه طويلتري همانند سازي ميشود و از محصول PCR اول كه بيشتر DNA هدف همانند سازي شده است يك واكنش ديگر با پرايمر هاي داخلي انجام ميگيرد. در اين روش اختصاصيت و حساسيت PCR بالا ميرود.
5- Long distance PCR : با اين روش قطعات ببيش از 20 kb همانند سازي ميشوند. براي انجام اين نوع PCR بايد DNA ژنومي از كيفيت بالايي برخوردار باشدو پرايمر ها به دقت طراحي شوند. براي اينكاراز klon taq استفاده ميشود.اين آنزيم نسخه اي از DNA poly است كه قسمت N ترمينال آن حذف شده است و با pfu poly به نسبت 180 به 1 مخلوط ميشود و فاقد 5` اگزو نوكلئازي است
كلونينگ با PCR :
مشكلات : اين جايگاه ممكن است روي قطعه همانند سازي شده نيز وجود داشته باشد.
گاهي اين جايگاه هنگام هضم شدن با آنزيم اشكال ايجاد ميكندكه بايد تعدادي باز به انتهاي 5` پراپمر اضافه شود.
استفاده از جايگاه دو آنريم متفاوت در هضم اشكال ايجاد ميكند.
2- Blunt end ligation : براي اينكار دو انتهاي قطعه همانند سازي شده بايد Polish شود اين كار توسط آنزيمهاي كلنو و T4 DNA polymerase ( پركردن قسمت over hang ) و يا آنزيم هاي S1 nuclease و Mong bean nuclease انجام ميشود
3- - T vector پلاسميد ناقل را با آنزيم EcoRV ويا هر آنزيم ديگركه DNA را بصورت Blunt برش ميدهد هضم ميكنند سپس توسط آنزيم Terminal transferase يا آنزيم Taq poly تعدادي T به انتهاهاي 3` رشته هاي پلاسميد خطي شده اضافه ميكنند .( لازم به ذكراست كه آنزيم Taq poly داراي خاصيت ترمينال ترانسفرازي است و تعدادي A به انتهاي 3` محصول آمپلي فاي شده اضافه ميكند ) . با انجام واكنش Ligation پلاسميد و محصول PCR را بهم متصل ميكنند.
4- توليد نيمه جايگاه شناسايي يك آنزيم برش دهنده در طرف 5` پرايمر. قطعات با T4 poly nucleotidkinase و ATP فسفريله ميشوند و سپس بهم متصل ميگردند ، بعد توسط يك آنزيم هضم شده و كلونينگ انجام ميگيرد .
ايجاد تغيير در محصول PCR :
ميتوان درسمت پايه5`` پرايمر يك پروموتور ويا جايگاه اتصال ريبوروم ( Ribosom Binding Site ) تعبيه نمود، همچنين ميتوان يك Non tanslated leader بين پروموتور و ژن قرار داد تا ناحيه براي شروع سنتز پروتئين مناسب باشد. اين كاربرد بنام Expression PCR گفته ميشود
20- Reverse Transcriptase PCR ( RT- PCR ):
تعدادي از آنزيم هاي پليمراز بجاي DNA از RNA بعنوان سوبسترا استفاده ميكنند واز روي RNA يك رشته DNA سنتز ميكنند. رشته جديد سنتز شده را cDNA ميگويند و واكنش بنام نسخه برداري معكوس Reverse transcription)) گفته ميشود. آنزيمهايي كه ار RNA بعنوان سوبسترا استفاده ميكنند Reverse trnscriptase معروف هستند. آنزيم Tth كه از باكتري ترموس ترموفيلوس بدست مي آيد در حضور يون منگنز فعاليت Reverse trnscriptase و در حضور يون منيزيوم فعاليت DNA polymerase دارد. آنزيم AMV كه از يك نوع ويروس پرندگان بنام Avian Myeloblastosis Virus trnscriptase استخراج ميگردد فعاليت RNase H هم دارد. دماي مطلوب فعاليت آن 42 درجه است و براي تهيه cDNA با طول كمتر از 500 b استفاده ميگردد. آنزيم MMLV ازيك نوع ويروس جوندگان بنام Mouse Molony Leukemia Virus استخراج ميشود و فعاليت RNase H آن كمتر از آنزيم قبلي ميباشد . در 37 درجه فعاليت ميكند . آنزيم Superscript آنزيم MMLV مهندسي شده است كه ژن قسمتي كه فعاليت RNAse H دارد را از آن حذف كرده اند و براي تهيه cDNA بزرگ استفاده ميگردد.
PCR با پرايمر هاي احتمالي (Degeneracy primer):
اين پرايمر ها بر اساس اطلاعات ترادف پروتئين يك ژن طراحي ميشوند و بر اساس اسيد هاي آمينه اي انجام ميگيرد كه داراي يك يا دو كدون باشند.پرايمر هايRAPD - بنام پرايمر هاي اختياري هم گفته ميشوند . اين پرايمر ها پلي مرفيسم را بدون شناخت توالي نوكلئوتيدي رديابي ميكنند
21– استخراج DNA از خون :
1- مقدار 500 ميكروليتر خون ( خون لخته نشده باشد – براي كارهاي PCR از EDTA بعنوان ضد انعقاد استفاده شود زيرا مواد ضد انعقاد ديگر مهار كننده آنزيم پليمراز ميباشند)
2- مقدار يك سي سي از باهر ليز كننده به آن اضافه كرده سانتريفيوژ نماييد
Lysis buffer = 0.33 M sucrose, 10mM Tris , 5mM MgCl2, 1% Triton X-100
3- مايع رويي را دور بريزيد و رسوب خاصل را چندين مرتبه در بافر فوق سوسپانسسيون نموده سانتريفيوژ كنيد تا رسوب بدست آمده شفبف گردد.
4- مقدار 400 ميكروليتر بافر ليز كننده به آن اضافه نموده سوسپانسيون كنيد و مدت 20 دقيقه بجوشانيد.
5- با فنل و كلرفرم آن را استخراج كنيد
6- با الكل آنرا روب داده و در 50 ميكروليتر آب حل كنيد
7- مقدار 2-5 ميكروليتر آن را براي واكنش PCR استفاده كنيد.
22- انچام واكنش PCR
توجه : آزمايش بايد در محلي بدون رفت و آمد انجام گيرد . سمپلر هاي مورد استفاده نبايد براي كارهاي ديگر استفاده شوند ، ظروف، لوله ها و سر سمپلر ها اتو كلاو شوند و هنگام كار از دستكش استفاده شود .
انجام واكنش PCR : مواد زير را داخل لوله مخصوص واكنش PCR بريزيد :
10mMdNTP 0.5 μl (0.1mM)
10x PCR buffer 5 μl
MgCl2 1.5 μl (1.5 mM)
Primer -1 20pmmol
Primer -2 20 pmol
Taq DNA polymerase 0.25 μl ( 1.25 U)
Template DNA 0.1- 1 μg
dH2O up to 50μl
لوله ها را داخل Block دستگاه ترموسايكلر قرار دهيدو دستگاه را روشن كنيد. (مقدار 100 μl روغن معدني روي واكنش بريزيد تا از بخار شدن مواد ممانعت بعمل آورد. لازم به ذكر است كه دستگاههاي ترموسايكلرجديد بصورت Heated lid ساخته شده اند يعني درب دستگاه كه روي لوله هاي واكنش قرار ميگيرد حدود 105 درجه گرم ميشود در نتيجه بالاي لوله گرمتر از پايين آن است و از بخار شدن مواد داخل لوله جلوگيري ميشود). پس از اتمام كار محصول آمپلي فاي شده را با آگارز 3 درصد الكتروفورز كنيد.
.23 - استخراج RNA از سرم:
از دستكش يكبار مصرف استفاده كنيد. تيپ و لوله هاي مورد استفاده را اتوكلاو شوند. در محيط كار و دستها RNase زياد است و RNA زود هيدروليز ميشود لوله , تيپ و مواد و لوازمي كه با RNA در تماس هستند نبايد با دست بدون دستكش تماس پيدا كنند.
1-مقدار 500 ميكروليتر از بافر RNXplus در لوله 5/1 سي سي بريزيد
2- مقدار 100 ميكروليتر از سرم به آن اضافه كنيد و درب لوله رابسته خوب محلوط كنيد
3- مقدار 100 ميكروليتر كلرفرم به آن اضافه كنيد
4- درب لوله رابسته و خوب بهم بزنيد و 2 دقيقه در هواي آزلد قرار دهيد
5- مدت 10 دقيقه با 10هزار دور در دقيقه سانتريفيوژ كنيد
6- مايع شفاف رويي را به لوله جديد منتقل كنيد.
7- دوبرابر حجم آن الكل مطلق اضافه كرده و 10 دقيقه سانتريفيوژ كرده مايع رويي را حذف كنيدو لوله را وارونه روي كاغذ جاذب الرطوبه قرار دهيد
8- مقدار 100 ميكروليتر الكل 75 درجه اضافه كرده سانتريفيوژ كنيد و سپس مايع رويي را خذف كرده و رسوب را خشك كنيد
9- رسوب را در 40 ميكروليتر DEPC treated water حل كنيد
24- انجام RT-PCR:
1- مقدار 10 ميكروليتر از RNA مرحله قبل را RT-PCR استفاده كنيد
RNA 10 μl
RT buffer 4 μl
RT enzyme 0.5 μl
RNasine 0.3 μl
dNTP 1 μl
primer (1&2) 20pmol
DEPC water up to 20 μl
قبل از اضافه كردن RNA مدت 10 دقيقه آن را در 70 درجه قرار دهيد تا از تشكيل لوپ جلوگيري شود. واكنش را يك ساعت در 42 رجه قرار دهيد و سپس 5 دقيقه در 94 قرار دهيد تاRT غير فعال شود وآنزيم پليمراز را مهار نكند. سپس به هر لوله مقدار 0.25 μl آنزيم پليمراز اضافه كنيد و با برنامه زير واكنش را 30 سيكل ادامه دهيد:
Denaturation 94o 30 sec
Annealing 55o 30 sec
Extension 72o 30 sec
براي واكنش Nested مقدار يك ميكروليتر از محصول PCR استفاده كنيد
Taq poly 0.25 μl
10 x PCR buffer 5 μl
Mg Cl2 1.5 μl
Primer (F&R) 3 μl
DNA 1 μl
dH2O up to 50 μl
واكنش را مختصري سانتريفيوژ كنيد وبا برنامه زير واكنش را 30 سيكل ادامه دهيد:
Denaturation 94o 30 sec
Annealing 57o 30 sec
Extension 72o 30 sec
در انتها مدت 5 دقيقه آن را دردماي 72 درجه قرار دهيد واكنش را با ژل آگارز 3 درصد الكتروفورز كنيد ( پرايمرها تعداد 250bp را از ناحيه 5`UTR ويروس هپاتيت C را آمپلي فاي ميكنند.
بايد ها و نبايد ها در PCR
2. اعمال قبل و بعد از PCR جدا از همديگر انجام گيرند
3. براي استفاده ازهر ماده از پيپت جدا گانه و اختصاصي استفاده كنيد
4. از پيپت هايي كه داراي plug هستند و يا از پيپت هاي يكبار مصرف استفاده كنيد
5. مواد ذخيره آزمايشگاه را تقسيم كرده و فريز كنيد و هرچند وقت صحت آنها راكنترل كنيد
6. هنگام استفاده هرلوله را spin كنيد تا موادي كه به اطراف درب لوله چسبيده اند رسوب كنند
7. چند ين كنترل منفي حين آزمايش Run كنيد
8. براي انحام آزمايشهاي تاييدي ازموادفريز شده استفاده كنيد
9. هيشه DNA آمپلي فاي شده را خارج از محل آماده كردن نمونه نگهداري كنيد
10. هنگام كار با PCR product مقداري از آن را جدا گانه نگهداريد
25- آنزيمها ي مورد استفاده درمهندسي ژنتيك
آنزيم هاابزار كار مهندسي ژنتيك در آزمايشگاه ميباشند و مخقق بدون آنها نميتواند كاري انجام دهد. در اين درس با انواع آنزيمها و طرز كار آنها آشناشده و مورد استفاده هر كدام را بررسي ميكنيم.
الف ) ليگازها :
اين آنزيمها اتصال فسفو دي استر بين تك رشته ها را روي رشته DNA تك رشته اي بر قرار ميكند. 5`-p يك رشته به 3`-OH رشته ديگر متصل ميشود.. كو فاكتور آنها ATP يا NAD است .
1- آنزيم DNA ligase : اين آنزيم از باكتري E.coli استخراج مي شود وكوفاكتور آن NAD است و اكثرا" اتصال بين دو DNA كه برش انتهايي ا نها بصورت Cohesive باشد را بر قرار ميكند و عده اي معتقدند كه اين آنزيم در فر قرار كردن اتصال بين دو DNA كه برش انتهايي آنها بصورت blunt باشد مشكل خواهد داشت .
2- آنزيم T4 DNA ligase : اين آنزيم از فاژ T4 استخراج ميشود وكوفاكتور آن ATP است . اين آنزيم اتصال هر دو انتهاي Cohesive و blunt را بر قرارميكند.
3- آنزيم T4 RNA ligase : اين آنزيم اتصال فسفو دي استربين 3`- OH و 5`-P مولكولهاي RNA را بر قرار ميكند . كوفاكتور آن ATP و Mg2+ است
ب) نوكلئاز ها :
اين آنزيم ها نوكلئو تيد هار ا تجزيه ميكنند.
4- آنزيم DNase I : يك اندو نوكلئاز است و dsDNA را به روش غير اختصاصي تجزيه ميكند . كوفاكتور آن Mg2+ ، + Ca2 و Mn2+ است .. اين آنزيم براي مقا صد زير استفاده ميشود:
- تجزيه غير اختصاصي DNA
- نشاندار كردن DNA
- ايجاد موتاسيون در ژن
5- نوكلئاز استافيلوكوكي : DNA و RNA را به روش غير اختصاصي تجزيه ميكند.
6- انزيم RNase A- اين آنزيم RNA تك رشته اي را هيدروليز ميكند.
7- آنزيم RNase H - اين آنزيم RNA را كه با DNA دوبلكس شده باشد راهيدروليزميكند و در سنتز رشته دوم cDNA كاربرد دارد.
8- آنزيم S1 Nuclease : اين آنزيم پلي نوكلئو تيد تك رشته اي را هيدروليز ميكند.
9- آنزيم Mong bean Nuclease – اين آنزيم شبيه S1 Nuclease عمل ميكند
10- آنزيم Exonuclease III (گزو نوكلئاز : (3`→ 5 فعاليت 3` فسفاتاز دارد
همچنين فعاليت شبيه RNase H هم دارددارد.
اين آنزيم اتصال فسفودي استرجايگاههاي آپوريني يا آپيريميديني را تجزيه ميكند
11- آنزيم Bal1 Nuclease : اين آنزيم ssDNA را به روش اگزو نوكلئازي يا اندو نوكلئازي تجزيه ميكند
ج ) آنزيم هاي محدود گر
د) متيلازها
12- فسفاتازها : اين آنزيمها فسفومونواسترازغيراختصاصي هستندكه درpH قليايي فعاليت ميكنند و درحضور پذيرنده هاي فسفات مانند اتانل آمين و Tris بعنوان فسفو ترانسفراز عمل ميكتتد .
فسفاتاز اتصال P--O را تجزيه ميكند ولي فسفوريلاز اتصال C--O را تجزيه ميكند
كاربرد فسفاتاز ها :
دفسفريلاسيون انتهاي فسفات اسيد نوكلئيك
وقتي با پروتئين يا اسيد نوكلئيك كنژوگه شوند بعنوان آنزيم رپورتر عمل ميكند
بعنوان leader در ترشح خارج سلولي و پروتئين هاي فيوژن عمل ميكند
13- آنزيمهاي T4 poly Nucleotid Kinase : اين آنزيمها فسفات γ را از NTP به مولكول پذيرنده (Aceptor) انتقال ميدهد و عبا رتند از:
پروتئين كيناز ها
كربو هيدرات كينازها
پلي نوكلئو تيد كينازها
آنزيم T4 poly nuleotide kinase براي فسفريله كردن يا نشاندار كردن 5`- OH استفاده ميشود ، همچنين براي اندازه گيري فعاليت اندونوكلئازي بكار ميرود.
ه ) DNA پليمرازها : گروهي از آنزيمها هستند هستند كه بكمك چهار نوكلئوتيد تري فسفات (dNTP) از روي رشته الگو يك رشته DNA سنتز ميكنند. اين آنزيمها براي فعاليت خود احتياج به پرايمر دارند. چنانچه داراي فعاليت 3→ 5`` اگزونوكلئازي باشند كار غلط گيري ( proof reading ) را هم انجام ميدهند
14- آنزيم DNA polymerase I :
اين آنزيم داراي فعاليت هاي زير است :
فعاليت 3→ 5`` اگرونوكلئازي
فعاليت 5`→ 3` DNA پليمرازي
فعاليت RNase H
فعاليت نوكلئازي
اين آنزيم براي كارهاي زير استفاده ميشود :
DNA LabelingNick translation
ايجاد انتهاهاي Blunt
15- آنزيم T4 DNA plymerase: يك آنزيم مولتي فانكشنال است وبراي كارهاي زيراستفاده ميشود.
داراي فعاليت DNA پليمرازل
فعاليت 3→ 5`` اگرونوكلئازي
براي PCR هم قابل استفاده است و در 37 درجه فعاليت ميكند
براي ايجاد انتهاهاي Blunt در dsDNA استفاده ميشود
فعاليت 5` نوكلئازي ندارد
16- آنزيم T7 DNA plymerase: اين آنزيم داراي فعاليت 3→ 5`` اگرونوكلئازي ميباشد
Sequenase نسخه اي از اين آنزيم است كه فاقد فعاليت 3→ 5`` اگرونوكلئازي ميباشد وبراي Sequencing استفاده ميشود
سرعت پليمريزاسيون اين آنزيم زياد است و براي سنتز رشته موتاژن در موتاژنزيس بكار ميرود
انتهاي 5`overhang را پر ميكند
17- آنزيم Taq DNA polymerase : اين آنزيم از باكتري گرمادوست ترموس اكواتيكوس استخراج ميشود
فعاليت 3→ 5 اگزو نوكلئازي ندارد
فعاليت 5` نوكلئازي دارد
فعاليت ترمينال ترانسفرازي دارد
آنزيمهاي نسخه بردار معكوس () : گروهي از DNA polymerase ها بنام RNA - dependent DNA polymerase يا Reverse Transcriptase گفته ميشوند اين آنزيمها RNA را الگو قرار داده و DNA مكمل آن را سنتز ميكند و عبارتند از:
18 - آنزيمAvian Myeloblastosis Virus ) AMV) : اين آنزيم از نوعي ويروس استخراج ميشود كه در پرندگان بيماري ايجاد ميكند .
اين آنزيم فعاليت RNase H هم دارد و براي سنتز رشته دوم cDNA استفاده ميشود
19- آنزيم Molony Murine Leukemia Virus ) MLMV) : اين آنزيم از نوعي ويروس استخراج ميشود كه در جوندگان بيماري ايجاد ميكند.
آين آنزيم فعاليت RNase H كمتري از آنزيم AMV دارد
20 – آنزيم ترمينال ترانسفراز :
اين آنزيم پليمريزاسيون dNTP هارا ازروي پايه 3`- OH انجام ميدهد . اين آنزيم براي اتصال نوكلئوتيد ها به انتهاي 3`- OH رشته DNA استفاده ميشود وبراي كلونينگ محصول PCR قابل استفاده است.
21- آ نزيم هاي RNA polymerase :
اين آنزيم ها اطلاعات ژنتيكي را از DNA به RNA منتقل ميكنندو عبارتند از
RNA پليمراز باكتريايي
T7 RNA پليمراز ، يك پروموتور قوي دارد
T3 RNA پليمراز
SP6 RNA پليمراز
22- آنزيم پليمراز A: :
اين آنزيم دم پلي A را سنتز ميكند ودر انتهاي3` رشته mRNA سلولهاي يوكاريوت غير هيستوني يافت ميشود.
اين آنزيم ميتواند RNA هاي polyA منفي را به polyA+ تبديل كندو آنها را بكمك oligo) dT) بعنوان پرايمر، به cDNA تبديل نمايد
23- آنزيم هاي رپورتر / ماركر:
ابن آنزيمها در كلونينگ استفاده ميشوند وچون سريعا" بيان ميشوند براي غربالگري كلني هاي باكتريايي مفيد ميباشند ( آنزيم بتا گالاكتوزيد ازبراي غربالگري توضيح داده ميشود)
ß گالاكتوزيداز - اتصالات گالاكتوزيدي را در اليگو ساكاريد ها هيدروليز ميكند
ß لاكتاماز - اتصالات آميدي را در حلقه ß لاكتام هيدروليز ميكند و پني سيلينوئيك اسيد و سفالوسپوروئيك اسيد توليد ميكنند كه از نظر بيولوژيك غير فعال هستند
-كلرامفنيكل استيل ترانسفراز ( CAT) - كلرامفيكل را غير فعال ميكنند
-لوسيفرازها - آنزيم هاي توليد كننده نور هستند.
آنزيم هاي ( برش دهنده) محدود گر ((Restriction Enzymes
اندو نوكلئازهاي محدود كننده گروهي از DNase ها ميباشند كه توالي نوكلئو تيدي خاصي را شناسايي ميكنند . اين آنزيم ها dsDNA را بصورت اختصاصي يا غير اختصاصي برش ميدهند. اين آنزيم ها اولين بار در دهه 1950 به عنوان سيستم هاي Resriction وModification ( باكتريها براي جلوگيري از حمله باكتريوفاژها و عناصر ژنتيك خارجي اين سيستم ها را بكار مي برند ) كشف شدند. باكتريها توسط سيستم R-M ميتوانند DNA اي را كه از خارج هنگام عفونت ،كنژو گاسيون و ترانسفكشن وارد آنها ميشود را تخريب كنند.سيستم R - M باكتريايي معادل سيستم ايمني پروكاريوتي ميباشد.سيستم هاي R-M كه در ميكرو ارگانيسمها ( باكتريها ) يافت ميشوند بسيار گوناگون ميباشند (حتي در داخل يك سلول ) . تعداد اين آنزيمها از 2100 فراتر رفته و 17 آنزيم نوع I ، 179 آنزيم نوع II و 4 آنزيم از نوع III وهمچنين مشخصات 190 متيل ترانسفراز تغيير دهند DNA تعيين و شناسايي شده است .
سيستم هايR-M دو فعاليت آنزيمي دارند:
الف ) يك اندونوكلئاز (R.ENase) با جايگاه اختصاصي كه مسئول هضم DNA خارجي ميباشد
ب) يك متيلاز تغيير دهنده DNA ( متيل ترانسفراز ) كه همان توالي ويژه را شناسايي ميكند . متيل ترانسفراز ( M.MTase) مسئول تغيير دادن وحفظDNA خودي از هضم شدن توسط R.ENase ميباشد. ممكن است فعاليت R وM در يك آنزيم (واحد) كه داراي چند زير واحد است قرار داشته و يا از نظر فيزيكي آنزيم هاي جداگانه باشند. علي رغم اينكه آنزيم هاي Restriction و Cognate Modifications سكانس مشابه را شناسايي ميكنند، ترادف اسيد آمينه آنها يكسان نيست، شايد اين آنزيم ها با مكانيسم متفاوت DNA هدف را شناسايي ميكنند .
انواع سيستم هاي آنزيمي Restriction - Modification
بر اساس تركيب آنزيم ، كوفاكتورهاي مورد نياز ، قرينگي توالي DNA اي كه شناسايي ميكنند و مشخصات هضم DNA به چهار نوع تقسيم ميشوند. لازم به ذكر است كه گروه متيل دهنده سوبسترا در تمام متيل ترانسفرازها AdoMet ميباشد.
آنزيم هاي R-M نوع I:
مجموعه چند آنزيمي متشكل از زير واحد هاي غير همسان بنام R,M و S ميباشند. زير واحد S اختصاصيت را براي MوR تعيين ميكند. كوفاكتور هاي اين آنزيمها ATP وMg2+ ميباشند و DNA اي كه تغييري در آن ايجاد نشده باشد را در محلي دور از جايگاه شناسايي و تا اندازه اي بصورت احتمالي ( Random ) برش ميدهند . هيدرو ليز ATP قسمتي از فعاليت Restriction اين آنزيم ها ميباشد و بطريق رقابتي با فعاليت اندون.كلئازي خودش جايگاه اختصاصي DNA هدف را متيله ميكنند.
آنزيم هاي R-m نوع II :
اين آنزيمها داراي فعاليت R.ENase و M.MTase جداگانه ميباشند . R.ENase ها دايمر بوده و زير واحد هاي آنها همسان است در صورتيكه M.MTase ها آنزيم هاي مونومر ميباشند . R.ENase هاي نوع II براي فعاليت خود فقط Mg2+ نياز دارند.
R.ENase ها و M.MTase ها تواليهاي اختصاصي نوكلئوتيدي مشابه را شناسايي ميكنند . اكثر R.ENase هاي نوع دوم DNA را ازمحل جايگاه شناسايي برش ميدهند ولي تعدادكمي از آنها كه نوع IIS گفته ميشوند DNA را در فاصله معيني از جايگاه شناسايي برش ميدهند .
آنزيم هاي R-M نوع III:
اين آنزيم ها متشكل از دو زير واحد غير همسان ميباشند كه براي فعاليت رستريكشن به ATPو Mg2+ نياز دارند. اين آنزيم هاتوالي هاي كوتاه غير پاليندرم را شناسايي كرده و DNA را از محلهاي معين (درصورتيكه درجايگاه شناسايي تغييري ايجادنشده باشد )برش ميدهند اين آنزيمها ATP را هيدروليز نميكنند و براي برش دادن DNA به AdoMet ( S-adenosylmethionine)نياز ندارند اما AdoMet موجب تحريك فعاليت اندو نوكلئازي آنهاميشود.
آنزيم هاي نوع IV :
اين آنزيمها از نظر تكاملي بين آنزيم هاي نوع IIوIII ميباشند. اين آنزيمها (مانند Eco571) متشكل از R.ENae و M.MTase جدا گانه ميباشند اما R.ENase كه يك آنزيم مونومر ميباشد فعاليت متيلازهم دارد. اين فعاليت متيلازي قدرت كافي ندارد تا DNA ميزبان را در In vivo محافظت كند.R.ENase و M.MTase توالي هاي ناقرينه را شناسايي ميكنند . R.ENase در حضور Mg2+ در فاصله معيني از جايگاه شناسايي, DNA هدف را برش ميدهد ( مثلا" Eco571 بفاصله 14/16 نوكلئو تيد از محل شناسايي اين كار را انجام ميدهد ) . فعاليت Restriction آنها توسط AdoMet تحريك ميشود.
سيستم هاي Modification يا Restriction مستقل
علاوه بر چهار سيستم آنزيمي كه بحث شد ، بعضي از باكتريها سيستم هاي ديگري مانند سيستم R مستقل از M و سيستم M مستقل از R دارند. يك مقوله اي از سيستم هاي R-M كه در تكنولوژي نو تركيب اهميت بخصوصي دارد Restriction وابسته به Methylation ) MDRS) ميباشد كه R.ENase ترجيحا" DNA را در جايگاههاي متيله برش ميدهد . MDRS متشكل از فرآورده هاي ژني است كه به چندين جايگاه مستقل در ژنوم E.coli K12 مانند mcrA , mcrB و mrr متصل ميباشند. جدول شماره 5- طبقه بندي سيستم هاي R-M
|
Feature |
نوع I |
نوع II |
نوع III |
نوع IV |
|
ساختمان فعال R-M |
يك آنزيم با سه زير واحد (R.M.S) |
انزيم هاي جدا گانه |
يك آنزيم با دو زير واحد |
آنزيم عاي مونومري جدا گانه |
|
كوفاكتورها |
ATPو mg 2+ |
mg 2+ |
ATPو mg 2+ |
(AdoMet) mg 2+ |
|
جايگاه شناسايي |
غير قرينه |
پاليندرم غير قرينهه |
عير قرينه |
غير قرينه |
|
هضم DNA |
فاصله متغير از هر طرف |
همان جايگاه به فاصله معيني از طره 3` |
25-27bpاز طرف 3` |
14bpاز طرف3` |
|
متيلاسيون |
دو رشته را بوسيله M متيله ميكند |
دو رشته يك متيل ترانسفراز روي هر رشته |
فقط يك رشته |
دو رشته |
كارگاه عملي – نظري كلونينگ , نشاندار كردن و هيبريديزاسيون اسيد هاي نوكلئيك(قسمت اول)
1- باكتري بعنوان ميزبان
2- پلاسميد به عنوان حامل ژن
3- آنزيم بعنوان وسيله برش و اتصال قطعات DNA به يكديگر
ژنوتيپ باكتريها
F or F`( Fertility)) : اپي زومي كه پيلي جنسي ايجاد ميكند . قسمت هايي از كروموزوم E.coli مانند اپرون ( Lac - pro A) روي F` قرار دارد. براي آلوده كردن باكتري به فاژ M13 يا فاژ هاي كمكي ( M13K07 , R408 ) فاكتور F` لازم ميباشد. اين فاكتور براي توليد DNA تك رشته اي از فاژ( فارميد ) مفيد ميباشد .
ْْْْْْْْْْْْْْْْْhsd R - اين موتاسيون موجب كاهش خاصيت رستريكشن EcoK ميشود اما روي متيلاسيون تاثيري ندارد. باكتري كه داراي اين موتاسيون باشد DNA خارجي بدون اينكه توسط R.ENase نوع I هضم شود وارد آن ميشود.
hsd S باكتري كه داراي اين موتاسيون باشد فعاليت متيلاسيون و رستريكشن آن كاهش ميابند.
mcr- ( Methyl Cytosine Restriction) ) اين موتاسيون اختصاصا" روي هر دويا يكي ازسيستم هاي McrA و McrB تاثير ميكند. ژن McrA روي كروموزوم E.coli قرارداشته وكد كننده فعاليت Restriction - Methylation اختصاصي براي CmCGG ميباشد . ناحيه mcrB در روي كروموزوم حاوي دو ژن mcrB و mcrC است . محصول اين دو ژن موجب تشكيل نوكلئاز وابسته به هترو دايمر GTP ميشود. سومين فعاليت mcr بنام mcrF گفته ميشود( به mrr مراجعه شود ). اگر در Dcm تغييري ايجاد شده باشد با +Mcr هضم نميشود.
mrr- قسمتي از سيستم رستريكشن وابسته به متيلاسيون(MDRS) است . اين ژن DNA حاوي آدنين هاي متيله شده را هضم ميكند. ژن mrr روي ترادف هاي حاوي سيتوزين متيله( CpG) فعاليت Restriction دارد. اين فعاليت اشاره به mcrF ميباشد. اگر درDam , Dcm يا EcoK تغييري ايجاد شود DNA با Mrr مثبت ويا هرسيستم رستريكشن ديگر E.coli K12 هضم نميشود.
proA - اين موتاسيون فعاليت گاما گلوتاميل فسفات ردكتاز را كاهش ميدهدو بيوسنتز پرولين ناقص ميگردد. باكتري موتانت براي رشد خود به اسيد آمينه پرولين نياز دارد.
proB - اين موتاسيون موجب كاهش فعاليت آنزيم گاما گلوتامات كيناز شده وبه نقص بيوسنتز پرولين منجر ميشود . باكتري موتانت براي رشد خودبه اسيد آمينه پرولين نياز دارد.
ara A - درژن سيستم ال- آرابينوز ايزومراز باكتري موتاسيون رخ داده است و باكتري موتانت نميتواند از آرابينوز استفاده كند.
araC - در ژن رگولاتوري ( پروتئين اكتيواتور/ رپرسور ) موتاسيون رخ داده است
galK - (galA) ژن ساختماني گالاكتوكيناز است. باكتريهاي موتانت نمي توانند گالاكتوز را مصرف كنند. اين موتانتها از گالاكتوز Expression Vector استفاده ميكنند.
LacY- اين موتاسيون موجب كاهش فعاليت گالاكتوز پرمئاز ( پروتئين M ) ميشود و باكتري نميتواند از لاكتوز استفاده كند .
recAI- اين موتاسيون موجب كاهش نو تركيبي هومولوگ و افزايش پايداري DNA كلون شده ميگردد. رشد اين سويه ها كند است و كارآيي ترانسفر ماسيون آنها پايين است.
rpsL- درژن زير واحد S12 ريبوزوم 30S موتاسيون ايجاد ميشود كه موجب مقاومت به استرپتومايسين ميشود. اين ژن بصورت StrA هم نوشته ميشود.
e14- اپي زوم شماره 14 حاوي ژن mcrA1 ميباشد و حذف آن موجب صفت McrA منفي در باكتري ميشود. حذف e14 منجر به آللهاي ديگر McrA ميشود.
supE44- اين موتاسيون موجب ميشود كه به جاي UAG كه كدون متوقف كننده است اسيد آمينه Gln در زنجيره پپتيدي قرار بگيرد. اين سويه ها حاوي ژن (gln V ) براي گلوتامين tRNA2 ميباشند كه براي رشد فاژها لازم ميباشند.
xyLA - موتاسيوني كه درD- گزيلوز ايزومراز ايجاد ميشودوباكتري موتانت نمي تواند گريلوز را مصرف كند.
LacIq- ايجاد موتاسيون در پروموتور i ( ژن رپرسور ) كه منجر به توليد 10 برابر رپرسور لاكتوز ميشود. اين موتان براي توقف بيان ( اكسپرس ) ژن از طريق پروموتور لاكتوز مفيد ميباشد.
amber codon - كدون UAG كه متوقف كننده ( Termination ) سنتز پروتئين است بنام آمبر كدون گفته ميشود.
Tn10- ترانسپوزوني كه موجب مقاومت به تتراسيكلين ميشود ( Tetr) .
endA -اين موتاسيون بصورت غير اختصاصي DNA endonuclease I را غير فعال ميكند . اين موتاسيون براي جدا كردن پلاسميد DNA مفيد ميباشد و كيفيت آن را بهبود مي بخشد.
gyrA96 -موتاسيون درژن زير واحدA آنزيم DNA gyrase ايجاد ميشود و باكتري به ناليديكسيك اسيد مقاوم ميگردد.
thi I(thi A)- موتاسيون درژن سنتز تيازول ايجادشده است وبراي رشد باكتري تيامين ( ويتامين B123 ) با منشاء خارجي نياز ميباشد.
supE - باكتري موتانت كدون UAG ( كدون ختم زنجيره ) را بعنوان كدون گلوتامين شناسايي ميكند. هنگام سنتز پروتئين وقتي با UAG مواجه شود اسيد آمينه گلوتامين وارد زنجيره پلي پپتيدي ميشود. كدون UAG بصورت عادي يك كدون متوقف كننده است .وكتور M13 براي كدون UAG درچندين ژن موجوددر ژنوم فاژ طراحي شده بود اين وكتورها فقط درسويه هاي آزمايشگاهي ذرات فاژ توليدميكردند. ژن آنها بطورصحيح در باكتري هاي سويه وحشي ترجمه نميشد (مانند باكتريهايي كه در روده زندگي ميكنند) وقادربه رشددرمحيط خارج از آزمايشگاه نبودند. اين ايده موجب جلوگيري از آزادكردن باكتريهاي مهندسي شده درمحيط گرديد.
F`[traD36]- ژن TraD درانتقالDNA ازطريق كنژوگاسيون دخالت دا (Fertility plasmid ) . ژن موتاسيون يافته ازكنژوگاسيون باكتري جلوگيري ميكند. پلاسميد هاي F` هنگام كنژوگاسيون از يك سلول به سلول ديگر منتقل ميشوند و يك پلاسميد مهندسي شده ميتواند به يك باكتري وحشي منتقل شود ( يعني باكتري ترانسفرم شده از پتري ديش خارج شود ). موتاسيون traD36 با كاهش پديده كنژوگاسيون يك نوع ايمني ميباشد كه از ايجاد حادثه جلوگيري ميكند.
F`[ proAB+] - اين موتاسيون موجب حذف ژنهاي بيوسنتز پرولين از باكتري شده واين ژنها در پلاسميد F` مستقر شده اند. بنا براين باكتري ميتواند پرولين را سنتز كند و احتياجي به اضافه نمودن پرولين به محيط كشت نميباشد ( تا زماني كه باكتري پلاسميد F` داشته باشد ). اگر باكتري پلاسميد F` خود را از دست بدهد ( مثلا" وقتي باكتري مدت زيادي در كشت آگار نگهداري شود ) به proAB منفي تبديل ميشود و باكتزي فقط در حضور پرولين رشد ميكند. اگرباكتري پلاسميد F` خود را از دست بدهد مدت زيادي نميتواند پيلي جنسي بسازد وبا فاژ M13 آلوده نميشود. بنا برا ين كشت باكتري در محيط minimum كه فاقد پرولين است انجام ميگيرد و به اين معني است كه فقط سلولهايي كه پلاسميد F` دارند و به M13 آلوده ميشوند ميتوانند زنده بمانند.
F`[LacZΔM15] - lacZ ژن بتا گالاكتوزيداز است . ΔM15 يعني اين نسخه از ژن LacZ فاقد كدونهاي 41-11 است . كدونهاي 41-11 مخصوص قسمتي از پروتئين LacZ هستند كه بنام پپتيد آلفا گفته ميشود. در غياب اين پپتيد آنزيم بتا گالاكتوزيداز ناقص است ( براي فعاليت آنزيم ، سلول بايد داراي ژن LacZ` باشد كه براي Missing peptide كد شود) . ژن LacZ` روي وكتور است وسلولهاي حاوي اين وكتورها ميتوانند ژن بتا گالاكتوزيداز كامل توليد كنند، بنا بر اين وكتور و سلول با همديگر در پديده Lac selection همكاري دارند.
نقش ژن LacZ- اين ژن ماركر انتخابي با كتري نوتركيب ميباشد و 140 اسيد آمينه آنزيم بتا گالاكتوزيدازباكتري E.coli را كد ميكند . ژن مذكورلاكتوزرا به گلوكز و گالاكتوز تبديل ميكند. پروتئيني كه توسط LacZكد ميشود به تنهايي براي تبديل لاكتوز به گلوكز و گالاكتوز كافي نيست و با يك قطعه ثانوي كه حاوي بقيه پروتئين بتا گالاكتوزيداز است كامل ميشود و آنزيم فعال توليد ميكند. اين پپتيد ثانويه توسط وكتور (براي ترانسفر ماسيون در آزمايش كلونينگ استفاده ميشود ) سنتز ميشود . واكنش ترانسفرماسيون باكتري با وكتورحاملLacZ` روي پليت آگار حاوي ) X-gal 5- برومو، 4 -كلرو، 3 - ايندوليل - بتا گالاكتوپيرانوزيد ) كه آنالوگ لاكتوز است پخش ميشود.
سيستم Lac selection بصورت زير كار ميكند: (Lac Operon)
سلولي كه حاوي وكتور كلونينگ با يك ژن LacZ` بكر ( دست نخورده ) است ميتواند بتا گالاكتوزيداز را سنتز كند اين آنزيم همراه با IPTG كه يك القا كننده است X-gal را به تركيب آبي رنگ برومو كلرو ايندوليل تبديل ميكند . اين سلولها كلني آبي رنگ (فقط حاوي پلاسميد هستند ) يا پلاك آبي رنگ ( حاوي فاژ ) توليد ميكنند. سلولهاي نوتركيبي كه ژن LacZ` آنها توسط Inserted DNA تخريب شده است نميتوانند بتا گالاكتو زيداز را سنتز كنند و كلني هاي سفيد يا پلاك هاي شفاف توليد ميكنند
- بافر هاي مورد نياز در آزمايشگاه بيولوژي مولكولي
اولين وظيفه فرد در آزمايشگاه مولكولار بيولوژي تهيه بافر هاي مورد نياز ميباشد. همه واكنشهاي بيولوژي در بافر مخصوص انجام ميشوند . معمولا"مواد اصلي بافر هارا در آزمايشگاه بصورت ذخيره ((Stock تهيه ميكنند و بافر هاي مورد نياز روزانه را از مواد ذخيره تهيه مينمايند . بعضي از مواد ذخيره و مولاريته آنهابه اين شرح است:
|
نام ماده |
مولاريته ( غلظت ) |
نام ماده |
مولاريته ( غلظت) |
|
Ammonium Acetate |
10M |
NaCl |
5M |
|
Tris pH ( 8, 7.5 , 7.4) |
1M |
CaCl2 |
1M |
|
NaOH |
5N |
EDTA |
0.5M |
|
Ethidium bromide |
1mg/ml |
SDS |
10% |
|
X-gal |
20mg/ml |
KAc |
5M |
|
IPTG |
200mg/ml |
NaAc |
3M |
براي تهيه محلول كار از مواد ذخيره از فرمول زير استفاده ميشود
M1V1=M2V2
M1 مولاريته ماده ذخيره V1حجم ماده ذخيره كه بايد براي بافر برداشته شود
M2مولاريته بافر موردنياز V2حجم بافرخواسته شده
مثلا" براي تهيه 50 ميلي ليتر TE بافر ( 10 ميلي مولار تريس و 1ميلي مولار EDTA ) چنين عمل ميكنيم :
50x10 = ?x1000, 500:1000= 0.5cc ( 500 μl) Tris
50x1 = ? x500 , 50:500 = 0.1cc ( 100μl ) EDTA
روش كار: 500μl از بافر تريس و 100μl از EDTA را درظرف مدرج ( لوله فالكن ) ريخته و حجم آن را به 50ml ميرسانيم .
3- تهيه محيط LB brothبراي كشت باكتري ( E.coli )
هدف از آموزش اين قسمت تهيه محيط مايع براي كشت با كتريهاي مورد استفاده در كار هاي بيولوژي مولكولي ميباشد
1- مواد زير را وزن كرده درون ارلن ماير ريخته و روي Stirrer مخلوط كنيد
تريپتون 5/2 گرم سديم كلرايد5/2 گرم
عصاره مخمر25/1 گرم آب مقطر250 ميلي ليتر
2- pH محيط را مساوي 5/7-7 تنظيم كنيد و سپس آن را اتوكلاو نماييد. مقدار لازم از آن را درون لوله مناسب ريخته ( در صورت نياز آنتي بيوتيك مناسب به آن اضافه نماييد ) و بصورت استريل ( كنار شعله ) يك كلني از سلول مورد نظر را درآن كشت داده ويك شب در 37 درجه آن را Shake كنيد.
4- تهيه آگار پليت
هدف از انجام اين كار تهيه محيط كشت جامد براي كشت باكتريها ميباشد. به محيط LB مقدار 5/1 درصد وزني پودر آگارآگار اضافه كرده وروي شعله قرار دهيد تا خوب بجوشد تا آگار كاملا" حل شده ومحلول شفاف بدست آيد سپس آن را اتوكلاو كنيد.مقدار 100μg / ml آمپي سيلين ( يا هر آنتي بيوتيك مناسب ديگر ) به آن اضافه كنيد.محيط را داخل چند پليت تقسيم كرده و در حرارت اتاق قرار داده تا آگار سفت شود سپس آنها را لاي كاغذ پيچيده و تا هنگام استفاده دربرودت 4 درجه نگهداري نماييد.
اطلاعات بيشتر
5- تهيه آنتي بيوتيك هاي مورد نياز كارگاه :
الف ) 500mg آمپي سيلين ( يك ويال) رادر 5 سي سي آب مقطراستريل حل كنيد غلظت آن100mg/ml است و هر ميكرو ليتر اين محلول حاوي 100μg آمپي سيلين است .آمپي سيلين با غلظت نهايي 100μg/ml مصرف ميشود. براي هر ميلي ليتر محيط كشت ( LB broth يا آگار پليت ) يك ميكروليتر از اين محلول استفاده ميشود.ب) تتراسيكلين ( ويال يا كپسول ) را با غلظت 25mg/ml در متانول حل كنيد و در لوله هاي يك ميلي ليتري تقسيم كرده فريزر كنيد. اين آنتي بيو تيك با غلظت نهايي 12.5μg/ml مصرف ميشود . براي هر ميلي ليتر محيط كشت( LB broth يا آگار پليت ) 0.5 ميكروليتر از اين محلول استفاده ميشود ( ميتوان محلول ذخيره تتراسيكلين را با غلظت 12.5mg/ml تهيه نمود سپس براي هر ميلي ليتر محيط يك ميكروليتر از ملول ذخيره اضافه نمود).
6- تهيه سلول پذيرا ( Competent cell )
هدف ازآموزش اين قسمت كسب مهارت براي تهيه سلول باكتري است بطوري كه طي پروسه اي كه روي آن انجام ميگيرد باكتري زنده بماند و پلاسميد (Circular DNA ) بتواند وارد آن شود. سرما وكلسيم كلرايد و متعاقب آن شوك گرمايي ( Heat shock) كه به سلول وارد ميشود تغييراتي در سطح سلول ايجاد ميشود وDNA پلاسميدي از طريق مناقذي كه روي سطح سلول ابجاد شده اند وارد آن ميگردد.
روش كار:
1- يك كلني از باكتري مورد نظر را در 3 ميلي ليتر محيط LB كشت داده ويك شب در 37 درجه روي شيكرانكوبه كنيد
2. يك حجم از كشت شبانه باكتري مورد نظر رادر ده حجم محيط LB كشت دهيد ( بنا به دلايلي كه بعدا" توضيح داده ميشود در اين كارگاهها از دو سويه HB101 و XL1-blue باكتري E.coli كه خصوصيات آنها نيز توضيح داده خواهد شد استفاده ميگردد)
3. مدت2.5ساعت در حرارت و rpm مناسب آن را Shake كنيد تا به OD600 = 0.6 برسد
4. مقدار يك ميلي ليتر آن را به لوله ميكروفيوژ انتقال دهيد
5. لوله حاوي سلول را ده دقيقه روي يخ قرار دهيد
6. مدت 5 دقيقه با 3500rpm سانتريفيوژ كنيد ( بهتر است در 4درجه انجام شود )
7. مايع رويي را خالي كرده ولوله را وارونه روي كاغذ جاذب الرطوبه قرار دهيد تا آب آن خارج شود
8. رسوب را بايك سوم حجم اوليه از محلول 100mM CaCl2 سوسپانسيون كنيد ( محلول نبايد ورتكس شود بلكه با ضربه انگشت آن را حل كنيد) و ده دقيقه روي يخ قرار دهيد
9. مانند مرحله 5 آن را سانتريفيوژ كرده محلو ل رويي را خالي كنيد
10. رسوب را با يك دوازده ونيم حجم اوليه محلول CaCl2 سوسپانسيون كنيد ( با ضربه دست )
آن را درون ميكروفيوژ هاي مناسب تقسيم كرده وبراي Transformation استفاده كنيد.
7- Transformation ( انتقال Plasmid DNA به باكتري زنده )
هدف از آموزش اين قسمت كسب مهارت لازم براي انتقال پلاسميد به سلول باكتري ميباشد . اين كار در بيولوژي مولكولي از اهميت بالايي بر خوردار است . براي كار كردن با ژنها بايد حتما" آن ژن در يك پلاسميد كلون شود و سپس پلاسميد نوتركيب (Recombinant plasmid) به سلول باكتري منقل شود تاپس از همانند سازي پلاسميد، ژن ( DNA ) كلون شده در پلاسميد نيز به تبع آن همانند سازي كند .
1. سلول پذيرا ( Competent cell ) را روي يخ قرار دهيد
2. Plasmid DNA مورد نظر را (5ng) به آن اضافه كرده خوب مخلوط كنيد ( بنا به دلايلي كه بعدا" توضيح داده ميشود دراين كارگاهها از دوپلاسميد pBR322 وBluescript كه خصوصيات آنها نيز توضيح داده خواهد شد استفاده ميگردد)
3. - لوله حاوي سلول پذيرا ( Competent cell ) را مدت 30 دقيقه روي يخ قرار دهيد
4.- واكنش را مدت 40 ثانيه در آب گرم 42 درجه قرار دهيد ( شوك گرما )
5.- مدت 2-1 دقيقه روي يخ قرار دهيد
6-. مقدار 200 ميكروليتر محيط LB ( بدون آنتي بيوتيك) به آن ضافه كرده و 60-40 دقيقه در 37 درجه انكوبه كنيد (باكتر ي E.coli هر 20 دقيقه يك نسل تكثير ميكند ودر اينجا به آن فرصت داده ميشود تا دو يا سه نسل تكثيرنمايد ) . .
7- بصورت زير روي آگار پليت پخش كنيد ( توجه : همه محتواي لوله را روي پليت خالي كرده و با Spreader آن را روي سطح آگار داخل پليت پخش كنيد):
پليتA : Competent cell+ plasmid DNA و آنتي بيوتيك مناسب
پليت: B Competent cell ( آگار حاوي آنتي بيوتيك است )
پليتC : Competent cell ( آگار فاقد آنتي بيوتيك است )
8.- پليت را به انكوباتور منقل كرده ودرب آنها را نيمه باز بگذاريد تاسطح پليتها خشك شود
9- . پليت هارا بصورت وارونه يك شب در حرارت 37 درجه انكوبه كنيد
10- اگر پروسه كار خوب انجام شده باشد بعد از 12 ساعت روي سطح پليت كلني ها ظاهر ميشوند
11- تفسير نتايج : اگر كار درست انجام شده باشد بايد روي پليت هاي A و C كلني باكتري رشد كرده باشد ولي روي پليت B كلني باكتري مشاهده نگردد رشد كلني باكتريايي روي پليت B به اين معني است كه باكتري مورد استفاده قبلا"با يك پلاسميد حاوي ژن مقاومت به آنتي بيوتيك ترانسفرم شده است در اينصورت كلني هايي كه روي پليت A ظاهر شده اند فاقد اعتبار ميباشند . اگر روي پليت C كلني با كتر ي رشد نكند به اين معني است كه Competent cell خراب بوده و باكتريها در طول پروسه كار مرده اندو نبايد انتظار داشته با شيم كه روي پليت A هم كلني باكتري رشد كند .كلني هاي باكتريايي كه روي پليت A رشد ميكنند كلني هاي حاوي پلاسميد مورد نظر هستند.
8- پلاسميد ها :
پلاسميد يك مولكول dsDNA حلقوي است كه وارد باكتري ميشود و همانند سازي آن داخل سيتوپلاسم باكتري وبطورمستقل از ژنوم باكتري انجام ميگيردوبه آساني هم از باكتري استخراج ميشود. هرپلاسميد داراي يك(ori) Origin of Replication ( منشاء همانند سازي ) است كه همانند سازي پلاسميد از آن نقطه شروع ميشود . قسمتي از ترادف پلاسميد كه براي تعدادي از Restriction enzyme ها فقط يك جايگاه برش دارد و اين آنزيم ها در قسمتهاي ديگرترادف پلاسميد جايگاه برش ندارند Multiple Cloning Site گفته ميشود وبراي تهيه Recombinant DNA از آن استفاده ميشود يعني مولكول DNA خارجي در اين قسمت كلون ( Clone ) ميگردد.تعدادي از پلاسميد ها در طبيعت يافت ميشوند (در باكتريها و مخمرها ) ولي در مقايسه با كروموزوم سلول ميزبان بسيار كوچكتر مي باشند و اغلب در خارج كروموزوم باكتري ديده ميشوند.( تعدادي از پلاسميد ها براي كارهاي خاصي در آزمايشگاه طراحي و ساخته شده اند ). اگر چه پلاسميدها براي زندگي سلول باكتري ضروري نيستند ولي حاوي ژن مقاومت به يك آنتي بيوتيك ميباشند كه براي كارهاي مهندسي ژنتيك لازم و ضروري است. بعضي از پلاسميدها تعداد زيادي كپي در داخل سلول باكتري دارند كه بنام High copy number plasmid گفته ميشوند(همانند سازي اين پلاسميد ها به كروموزوم باكتري بستگي ندارد) و Relaxed Plasmid هم گفته ميشوند . تعداد ديگري ازپلاسميدها تعداد كپي محدود تري در داخل سلول باكتري ايجاد ميكنند كه بنام Low copy number plasmid معروف هستند و باكتري شديدا" Replication آنها را كنترل ميكند و به Stringent Plasmid هم معروف ميباشند. با تغييراتي كه در Sequence پلاسميد ها ايجاد ميكنند صفات خاصي در آنها بوجود مي آيد كه در مهندسي ژننتيك استفاده ميشوند .چنانچه ori دو پلاسميد از يك منشا باشند هنگام ترانسفورماسيون با هم رقابت كرده و فقط يكي از آنها وارد باكتري ميشود اما اگر منشا ori انها با هم اختلاف داشته باشد هنگام ترانسفور ماسيون دو پلاسميد نيز ميتوانند وارد يك سلول بشوند ( اين مسئله در كارهاي كلونينگ اهميت خاصي دارد ).كارهاي عملي در اين كار گاههاي آموزشي با دو پلاسميد انجام ميگيرد:
الف) پلاسميد pBR322 اين پلاسميد توسط فرانسيسكو بوليوار و همكاران طراحي و ساخته شده است و همه خصوصيات يك پلاسميد خوب را دارا ميباشد . وزن مولكولي پايين (4363bp ) دارد و براي تعدادي از Restiction Enzyme ها فقط يك جايگاه برش دارد . اين پلاسميد داراي دوژن مقاومت به آمپي سيلين و تترا سيكلين ميباشد و هنگاميكه وارد باكتري ميشود باكتري نسبت به دو آنتي بيوتيك مذكور مقاوم ميشود.قسمتي از ترادف اين پلاسميد (705bp ) را كه براي ادامه زندگي آن ضروري نميباشد با آنزيم HaeII برش داده و حذف كرده اند واز آن پلاسميدي بنام pT153 ساخته اند كه همانند سازي آن 5-3 برابر پلاسميد PBR322 ميباشد ب) پلاسميد Bluescript sk همانطور كه در شكل ديده ميشود ترادف ژن Lacz` ( قسمتN terminal ژن β-galactosidase ) بطوري روي پلاسميد تعبيه شده است كه Multiple Cloning Site اين پلاسميد قسمتي از ترادف ژن مذكور ميباشد. (جايگاه برش آنزيمها درقسمت هاي مختلف ترادف پلاسميد PBR322 قرار دارند ) كه درمهندسي ژنتيك براي انتخاب كلني هاي سفيد باكتريايي حاوي Recombinant DNA استفاده ميشود. در دو طرف MCS اين پلاسميد پروموتور فاژهاي T3 و T7 قرار داشته كه ژن كلون شده را بر حسب Orientation آن ميتوان توسط هر يك از پروموتورهاي مذكور بيان ( Express) نمود . اين پلاسميد داراي دو Original of Replication است كه يكي از باكتري E.coli و ديگري از فاژ F1 گرفته شده است كه به اين علت فاژميد هم گفته ميشود.. اين پلاسميد براي توليد ssDNA هنگام Sequencing يك قطعه DNA هم قابل استفاده ميباشد. در Bluescript sk (M13) جايگاه آنزيم SacI بلافاصله در Downstream پروموتور فاژ T7 و جايگاه آنزيم KpnI بلافاصله در Downstream پروموتورفاژ T3 قراردارند. پلي كلونينگ سايت Bluescript ks (M13)) در orientation مخالف پلاسميد Bluescript sk (M13-) قرار گرفته است.
9- استخراج پلاسميد در مقياس كم ( Minipreparation )
هدف از آموزش اين قسمت كسب مهارت براي استحراج پلاسميد از داخل باكتري ميباشد و همانطوريكه قبلا" گفته شد اين كار در مهندسي ژنتيك و بيولوژي مولكولي از اهميت بالايي برخوردار است. استخراج Plasmid DNA با روش آلكالين وبه صورت Miniprep در موارد Colony screening استفاده ميشود و براي كارهاي حساس تر پلاسميد بصورت Large scale استخراج ميگردد. ( دراين كارگاه فقط Miniprep انحام ميشود). براي استخراج پلاسميد سلول باكتري با محلول GTE ( گلوكز، تريس ، EDTA ) ليز شده و محتويات آن آزاد ميشوند . Genomic DNA و پروتئينهاي سلولي را با محلول قليا ( NaOHو SDS ) دناتوره كرده(در اين روش DNA سوپر كويل دناتوره نميشود) و بلافاصله با اسيدي كردن محيط و ايحاد سرما Genomic DNA و پروتئينهارا رسوب ميدهيم. در اين پروسه Plasmid DNA بصورت محلول باقي ميماندكه توسط سانتريفيوژآنرا از فازآلي ( پروتئين ها ) جدا ميكنيم .
روش كار:
1- مقدار 5/1 ميلي ليتر كشت شبانه ( باكتري حاوي پلاسميد) را بمدت5 دقيقه با 4500rpm سانتريفيوژ كنيد . عده اي معتقدند چنانچه رسوب را با محلول STE سوسپانسيون كرده و دوباره سانتريفيوژ كنيم ( شستشو دهيم ) واكنش هاي آنزيمي بهتر انجام ميشود
[ STE = 100mM NaCl , 10 mMTris ( pH=8) , 1mM EDTA (pH=8)]
2- رسوب را در 100μl محلول I سوسپانسيون ( محلول روي يخ سرد شده باشد )
محلول I : 50mM glucose , 25mm Tris , 10 mM EDTA (pH 8)
3-. ميكرو فيوژ حاوي واكنش را مدت 10-5 دقيقه در هواي اتاق قرار دهيد
4.- مقدار 200μl از محلول II تازه تهيه شده به لوله واكنش اضافه كرده ، درب لوله را ببنديد و با وارونه كردن آن محلول را خوب مخلوط كنيدو 5 دقيقه روي يخ قرار دهيد.
محلول II : 0.2N NaOH, 1% SDS
5- مقدار 150μl از محلول III (سردشده روي يخ ) به لوله واكنش اضافه كنيد . درب لوله را بسته و به آهستگي 10 دفعه آن را وارونه كنيد ومدت 15 دقيقه روي يخ قرار دهيد.
محلول III : 60 ميلي ليتر 5M AcoK , اسيد استيك 5/11 ميلي لير, آب مقطر5/28 ميلي ليتر ( اين محلول داراي 3مولار پتاسيم و 5 مولار استات است ) وقتي اين محلولبه واكنش اضافه ميشود بايد يك رسوب سفيد رنگ در داخل لوله پديدار گردد .
6- مدت 20 دقيقه با 12000xg آن را سانتريفيوژ كنيد
7- محلول رويي (تقريبا" 450 ميكرو ليتر ) را به لوله ديگر انتقال دهيد( حاوي پلاسميد ميباشد)
8- سديم استات با غلظت نهايي 0.3M به آن اضافه كنيد ( عده اي اين مرحله راضروري نميدانند )
9- دو برابر حجم آن اتانل مطلق سرد اضافه كنيد
10- مدت 15 دقيقه آن را سانتريفيوژ كنيد تا plasmid DNA رسوب كند . الكل را حذف كنيد ( حتما" لوله را وارونه روي كاغذجاذب الرطوبه قرار دهيد ).
11- روي رسوب الكل 70درجه ( 100 ميكرليتر) اضافه كنيد و آن را ورتكس كنيد بطوريكه pellet كنده شده و در الكل شناور شود
12- مدت 3دقيقه در حرارت اتاق آن را سانتريفيوژ كنيد ( شستشو دادن با الكل )
13- الكل را خالي كرده ورسوب را در حرارت 70 درجه خشك كنيد .
14- رسوب را در 50 ميكروليتر آب يا بافر TE )) حل كنيد
TE= 10mM Tris pH 8 , 1mM EDTA pH 8
10- استخراج پلاسميد در مقياس بالا (Large scale Plasmid DNA preparation) :
1. مقدار 500 ميلي ليتركشت شبانه را با 4000g به مدت 10 دقيقه سانتريفيوژ كنيد. مايع رويي را خالي كرده و لوله را وارونه روي كاغذ خشك كن قرار دهيد تا درناژشود. رسوب را با STE سوسپانسيون نموده و دو بار ه سانتزيفيوژ كنيدو سپس لوله وارونه قرار داده تا درناژ شود.
[ STE = 100mM NaCl , 10 mMTris- base ( pH=8) , 1mM EDTA (pH=8)]
2. رسوب را در10 ميلي ليتر محلول I حل كنيد.
Solution I: 50 mM glucose , 25 mM Tris ( pH 8), 10 mMEDTA
3. آن را خوب سوسپانسيون كنيد و 10 دقيقه در هواي اتاق قرار دهيد
4. مقدار 20 ميلي ليتر از محلول II تازه تهيه شده به آن اضافه كرده درب لوله را ببنديد و با وارونه كرده آن محلول را خوب مخلوط كنيدو 5 دقيقه روي يخ قرار دهيدمحلول
Solution II: 0.2N NaOH, 1% SDS
5. مقدار 15 ميلي ليتر از محلول III به به واكنش اضافه كنيد و15 دقيقه لوله را روي يخ قرار دهيد
Solution III : 60 ml of 5M ACOK , 11.5 ml of Acetic Acid , 28.5 ml of ddH2O
( اين محلول داراي 3مولار پتاسيم و 5 مولار استات است ) وقتي اين محلول اضافه ميشود بايد يك رسوب سفيد رنگ پديدار گردد ( ميتوان باپروخالي كردن محلول توسط پيپت پروتئينها را شكست )
6. مدت 20 دقيقه با 20000xg آن را سانتريفيوژ كنيد
7. محلول رويي ر ا (تقريبا" مقدار 15 ميلي ليتر) به لوله جديد انتقال دهيد
8. مقدار0.6 حجم اوليه ( 12 ميلي ليتر ) ايزو پروپانل به آن اضافه كنيد وخوب مخلوط كرده و 15 دقيقه در حرارت اتاق قرار دهيد
9. درحرارت اتاق و مدت 15 دقيقه آن را سانتريفيوژ كنيد تا plasmid DNA در ته لوله رسوب كند ( توجه : اگر در 4 درجه انجام شود نمك ها رسوب ميكنند )
مايع رويي را حذف كنيد ( حتما" لوله را وارونه قرار دهيد ) و رسوب رابا الكل 70درجه و در حرارت اتاق شستشو دهيد ( مقدار 10 ميلي ليتر الكل 70 درجه به هر لوله اضافه كرده و با 17000g بمد ت 10 دقيقه سانتريفيوژ كنيد ). الكل را خالي كرده و آن را در حرارت 70 درجه خشك كنيد. رسوب را با 8 ميلي ليتر آب يا بافر TE حل كنيد .
11- هضمDNA باآنزيم هاي محدودكننده (restriction digestion ) ( كارها روي يخ انجام گيرد):
هر يك ازآنزيمهاي محدودكننده ( برش دهنده ) ترادف خاصي را روي رشته DNA شناسايي كرده ودورشته DNA را برش ميدهند . آنزيمها ي محدود گر در مهندسي ژنتيك ازاهميت بالايي بر خوردارند و در حقيقت شبيه چاقوي جراحي براي مهندسان ژنتيك ميباشند ( به جدول آنزيمها مراجعه شود ).
1-. مقدار پلاسميد را تخمين بزنيد ( يك واحد آنزيم مقدار يك ميكرو گرم DNA را در حرارت مناسب در مدت يك ساعت هضم ميكندويك ميكروگرم DNA را در يك واكنشي به حجم 20-30 ميكروليتر با آنزيم هضم كنيد)
2- باآب مقطر حجم آن رابه 18 ميكرليتر ( يا هرحجم مورد نظر) برسانيد
3- لوله واكنش را spin كنيد
4- مقدار 2ميكروليتر از 10x Reaction buffer به واكنش اضافه كنيد(غلظت نهايي بافر 1x باشد)
5- با استفاده از سرسمپلر استريل و سرد مقدار آنزيم مورد نظر را به واكنش اضافه نماييد ( اين مقدار آنزيم بايد در حجم نهايي در نظر گرفته شده باشد).
6- با ضربه نوك انگشت واكنش را مخلوط كرده و سپس spin نماييد.
7- مدت يك تا دوساعت در حرارت مناسب انكوبه كنيد
مقدار 15 ميكروليتر از واكنش را با 3 ميكرو ليتر loading buffer مخلوط كرده و روي ژل آگارز با درصد مناسب الكتروفورزنماييد ( از Intact plasmid DNA بعنوان كنترل واكنش روي ژل دركنارنمونه الكتروفورز شود توجه : اگر DNA هضم نشده است آن را P.C.I extraction نموده و با الكل رسوب داده وواكنش را تكرار كنيد ).
الكتروفورز DNA
جدول شماره 2 - قدرت جداسازي مولكولهاي DNA توسط غلظتهاي مختلف آگارز
|
درصد آگارز |
قدرت جداسازي DNA |
|
3 |
100-1000bp |
|
2 |
200- 1500bp |
|
1.5 |
300- 3000bp |
|
1 |
500 – 5000bp |
|
0.8 |
1000- 7000bp |
|
0.6 |
10 – 30kbp |
|
0.4 |
30 – 50kbp |
1- قالب ژل را آماده كنيد
2- شانه اي با دنده هاي مناسب روي قالب قراردهيد
3- حجم ژل مورد نظر را تعيين كنيد ( اندازه گيري طول وعرض قالب و قطر ژل مورد نظر )
4- پودر آگارز را وزن كنيد
5- آگارز را در بافر 1xTBE حل كنيد و آن را حرارت داده تا خوب بجوشد و قبل از خنك شدن به ازاي هر 10 ميلي ليترژل يك ميكروليتر از محلول 10mg/ml اتيديوم برومايد اضافه كنيد سپس آن را خوب مخلوط كرده و داخل قالب بريزيد
1xTBE= 89mM Tris base, pH8, 89mM boric acid, 2.5mM EDTA, pH8
6- بعد از بسته شدن ژل آن را از قالب خارج كرده و داخل تانك الكتروفورز قرار دهيد
7- هر 5 ميكروليترنمونه را با يك ميكروليتر بافر نمونه مخلوط كرده و داخل چاهك هاي ژل بريزيد (اين بافر حاوي 50درصد ماده سنگين كننده مانند گليسرين يا ساكارز است كه نمونه بخوبي داخل چاهك ريخته ميشود و همچنين حاوي0.2 درصد از يك رنگ نشانه مانند بروموفنل بلو يا گزيلن سيانول ميباشد . رنگ برومو فنل بلودر ژل 1.5 درصد همراه با 500bp و درژل 0.8 درصد با 1000bpحركت ميكند )
جدول شماره 3 : مقايسه حركت رنگ نشانه و ماركر DNA روي ژل آگارز
|
درصد ژل آگارز |
حركت تقريبي رنگ نشانه در مقايسه با ماركر |
حركت تقريبي DNA در مقايسه با ماركر | |
|
برومو فنل بلو |
Xylen cyanol | ||
|
0.4 |
500bp |
3 kbp |
2.5 kbp |
|
0.8 |
500 bp |
3 kbp |
1kbp |
|
1 |
500bp |
3 kbp |
500 bp |
|
1.25 |
500 bp |
3 kbp |
250bp |
|
1.5 |
100bp |
1kbp |
250bp |
|
2 |
100bp |
1kbp |
100bp |
|
درصد ژل پلي آكريلاميد |
حركت تقريبي رنگ نشانه در مقايسه با ماركر |
حركت تقريبي DNA در مقايسه با ماركر | |
|
برومو فنل بلو |
Xylen cyanol | ||
|
3.5 |
100bp |
500 bp |
100 bp |
|
5 |
75bp |
250bp |
75bp |
|
8 |
50bp |
250bp |
50 bp |
|
12 |
25bp |
75bp |
25bp |
|
15 |
10bp |
60bp |
25bp |
|
20 |
10bp |
60bp |
5bp |
9- DNA داراي شارژ منفي است و براي الكتروفورز شدن بايد نمونه بطرف قطب منفي با شد تا هنگام الكترو فورز به سمت قطب مثبت حركت كند
10- تانك الكتروفورز را به منبع تغذيه وصل كنيد
11- پيچ Current را روي ماكزيمم قرار داده و ولتاژ را تنظيم كنيد (10V/centimetr )
وقتي رنگ نشانه سه چهارم طول ژل را طي كرد ژل را از تانك خارج كرده بانورUV نتيجه را مشاهده كنيد .
13- تهيه ژل پلي آكريلاميد :
قدرت تفكيك سازي اين ژل بسيار بالا است و قطعات DNA با طول كم نيز در اين ژل قابل رويت ميباشند و بسته به در صد ژل حتي اختلاف يك باز را ميتوان در قطعات DNA الكتروفورز شده مشاهده نمود .
1. شيشه ها، شانه ( Comb ) و فضاسازها ( Spacer) را با آب و دترژان خوب بشوييد و سپس با الكل 70 در صد آنهارا تميز كنيد تا چربي دستها روي آنها باقي نماند ( بجاي پنبه از گاز استفاده كنيد زيرا پرزهاي پنبه روي شيشه باقي ميمانند ) .
2. قالب ژل را آماده كرده و جهت جلوگيري از نشت ژل ازداخل آن اطراف آن را با آگارز 1در صد به دقت درز گيري (Seal) نماييد .
3. حجم قالب (cast ) را ( حجم ژل) با اندازه گيري طول وعرض قالب مشخص نماييد ( قطر ژل بسته به قطر فضا سازها يك و يا نيم ميليمتر است )
4. بسته به در صدژل مورد نظر مقدار آكريلاميد و بيس آكريلاميد لازم را (بصورت مخلوط 29 در صد آكريلاميد و 1درصد بيس آكريلاميد تهيه و در يحچال نگهداري ميشود) داخل لوله تميز بريزيد
5. از بافر 10xTBE با غلظت نهايي 1x اضافه كنيد وبا آب مقطر به حجم مورد نظر برسانيد
6. مقدار 100 ميكرو ليتر آمونيوم پرسولفات 10درصد و 3 ميكرو ليتر محلول TEMED به آن اضافه كنيد ( توجه داشته باشيد كه مونومرهاي اكريلاميد وبيس آكريلاميد در حال پليمريزه شدن هستند)
7. به آرامي محلول را در قالب از قبل آماده شده بريزيد و مراقب باشيد كه حباب هوا وارد آن نشود زيرا حباب هوا مانع انجام الكترو فورز ميشود .
8. به آرامي شانه را داخل قالب قرار دهيد تا بعد از پليمريزه شده ژل ، درون آن چاهك هايي جهت نمونه گذاري ايجاد شود
9. بعد از پليمريزه شده به آرامي شانه را درآورده و چاهك هارا با آب بشوييد تا اگر مونومرهاي پليمريزه نشده هنوزوجود دارد شسته شوند
10. گيره پاييني را باز كنيد وفضاساز پاييني را از قالب خارج كنيد
11. گيره هاي طرفين را باز كرده و دستگاه را طوري داخل تانك مخصوص قرار دهيدكه قسمت بريده شده شيشه رويي ( Notch ) به سمت محفظه بالايي ( تانك فوقاني) قرار گيرد
12. شيشه هاي دستگاه ( قالب) را با نصب گيره ها ي مخصوص به طرفين تانك , ثابت نماييد
13. محفظه هاي بالا و پايين تانك را با 1xTBE buffer پرنماييد بطوريكه بافر روي چاهك ها را بپوشاند. چنانچه بين دوشيشه قالب ( محل خارج كردن فضاساز پاييني ) كه در محفظه پاييني تانك قرار دارد حباب هوا مشاهده ميشود با يك سرنگ پر از بافر كه دار اي سر سوزن كج است حباب هوا را بيرون كنيد تا مانع جريان الكتريكي نشود و الكترو فورز بخوبي انجام گيرد
14. محصول PCR را ( همانند الكتروفورز با ژل آگارز) با بافر نمونه گذاري مخلوط كرده و توسط سرنگ هاميلتون نمونه گذاري را روي ژل انجام دهيد
15. محفظه بالايي تانك را به قطب منفي ومحفظه پاييني را به قطب مثبت منبع تغذيه وصل كرده و الكتروفورز كنيد
16. بعد ازخاتمه الكتروفورز ( وقتي رنگ نشانه به پايين ژل رسيد) جريان برق را قطع كرده و شيشه را از تانك جدا كنيد. با يك تيغه فلزي نازك به آرامي بين دو شيشه فشار آوريد تا دو شيشه از هم جداشوند . ژل به يكي از دو شيشه مي چسبد . به آرامي آنرا داخل آب حاوي 10 ميكروگرم درهرميلي ليتر اتيديوم برومايد قرار دهيد تا بعد از چند دقيقه ژل رنگ شود .
ژل را روي دستگاه UV Transilluminator قرار داده و نوارهاي هاي DNA را مشاهده كنيد.
14-P.C.I extraction
هدف از اين كار حذف پروتئنهاي موجود در محلول حاوي DNA ميباشد . فنل ( equilibrated phenol با pH بيشتر از 7.5) موجب دناتوده شدن پروتئنها ميشود و كلرفرم نيز بقاياي فنل را از محلول واكنش حذف ميكند .
1- هم حجم محلول حاوي DNA فنل اضافه كنيد و مدت 5 دقيقه سانتريفيوژ كنيد ( فنل متعادل شده به علت 8-hydroxyquinoline ماده آنتي اكسيدان ) زرد رنگ است و روي آن با يك لايه بافر تريس پوشانيده شده است هنگام استفاده از فنل زير بافر استفاده كنيد)
2- مايع رويي ( مايع شفاف ) را به لوله تميز ديگري انتقال دهيد ( فنل در زير قرار ميگيرد و پروتئنهاي دناتوره شده بين دو لايه قرار ميگيرند).
3- هم حجم آن از مخلوط كلرفرم - ايزو آميل الكل ( 1:24) اضافه كنيد( ايزو آميل الكل يك ماده Anti foam است و مانع كف كردن محلول ميشود ) و مانند مرحله قبل ادامه دهيد
4- مايع رويي را به لوله تميز ديگر انتقال دهيد ويا مانند روش زير ميتوان عمل نمود:
5- مخلوط فنل - كلرفروم - ايزو آميل الكل (به نسبت 1:24:25) تهيه كنيد
6- حجم مساوي از مخلوط P.C.I به محلول حاوي DNA اضافه كنيد و 5 دقيقه سانتريفيوژ كنيد
7- محلول رويي را به لوله تميز ديگر انتقال دهيد و هم حجم آن C.I اضافه كنيد و 5 دقيقه سانتريفيوژ كنيد
8- محلول رويي را به لوله تميز ديگر انتقال دهيد (محلول حاوي DNA است )
9- اگر هنوز بوي فنل از آن متصاعد ميشود مرحله 7 را تكرار كنيد.
10- DNA را با الكل رسوب دهيد( تغليظ DNA ).
15- رسوب DNA با الكل ( تغليظ DNA )
هدف از اين كار احياي DNA بعد از استخراج و يا بعد از P.C.I. extraction ميباشد و در مواقعي نيز كه DNA خيلي رقيق باشد بدينوسيله آن را تغليظ ميكنيم.
1- حجم محلول حاوي DNA را تعيين كنيد ( تخمين بزنيد )
2- استات سديم با غلظت نهايي 0.3M به آن اضافه كنيد .
3.- مقدار دوبرابر حجم آن الكل مطلق سرد ( 20- يا 70-) اضافه كنيد
4- مدت 10 دقيقه با 12000xg سانتريفيوژ كنيد . رسوب سفيد رنگ را ميتوان در ته لوله مشاهده نمود ( لازم به ذكر است كه DNA خالص تقريبا" بيرنگ است ).
5- . با وارونه كردن لوله , مايع رويي را حذف كنيد و لوله را بصورت وارونه روي كاغذ جاذب الرطوبه قرار دهيد تا آب آن حذف شود .
6- مقدار 100 ميكرو ليتر الكل 70 درجه به آن اضافه كنيد و آن را ورتكس كنيد تا رسوب DNA از ته لوله كنده شده و شناور گردد ( اگر DNA بزرگ با شد ورتكس موجب شكسته شدن آن ميشود) .
7- مدت 5 دقيقه آن را سانتريفيوژ كنيد ( شستن DNA)
الكل را خالي كرده و DNA را در 70 درجه خشك كنيد ( چنانچه در اين مرحله DNA خوب خشك نشود والكل همراه آب داخل آن بماند بعدا" براي كار كردن با آنزيمها دچار اشكال ميشويد). DNA را در مقدار بافر ( آب ) دلخواه حل كنيد .
16- احياي DNA از ژل آگارز
هدف از اين قسمت كسب مهارت براي احياي DNA از روي ژل آگارز ميباشد. اين تكنيك در بيولوژي مولكولي و مهندسي ژنتيك اهميت زيادي دارد و در كلونينگ و انتقال ژن استفاده ميشود لازم به ذكر است كه از طرق مختلف اين كار انجام ميشود مانندFreez- Squeez, paper elution, Electero elution , و... ولي فقط دو روش توضيح داده ميشود :
الف ) احياي DNA ازروي ژل توسط كاغذ ( paper elution )
1- DNA مورد نظر رابا ژل آگارز 1-2درصد الكتروفورز كنيد و اجازه دهيد تانوارهاي هاي DNA بخوبي از همديگر تفكيك شوند
2- با نور UV ( طول موج 254 نانومتر موجب آسب DNA ميشود و بهتر است از طول موج 312 نانومتر استفاده گردد ) نوارهاي مورد نظر را مشاهده كرده و جلو آن را با تيغ اسكالپر تميز علامت گذاري كنيد
3- در جلو band يك حفره ايجاد كنيد ( عرض حفره مساوي عرض band باشد )
4- يك قطعه كاغذ واتمن و يك قطعه كيسه دياليز ( dialysis tubing ) به اندازه حفره ببريد
5.- كاغذ واتمن را در جلو حفره و كيسه دياليز را در پشت كاغذ قرار دهيد
6- الكترو فورز را ادامه دهيد ( بافر تانك را كمتر كنيد بطوري كه روي ژل را نپوشاند )
7- وقتي DNA وارد كاغذ شد الكترو فورز را متوقف كنيد
8.- كاغذ را جمع كرده و به لوله 5/ ميلي ليتري كه انتهاي آن سواخ شده است انتقال دهيد
9. توسط سمپلر مقدار مناسب با فر
(150mM NaCl , 20mM Tris (pH=7.5) 1mM EDTA , 5%SDS) روي كاغذ بريزيد
10- لوله مذكور را داخل يك لوله بزرگتر قرار دهيد ( 5/1 ميلي ليتر ) ، سانتريفيوژ كنيد تا مايع از ته لوله كوچك به داخل لوله بزرگتر حركت كند ، چند دفعه اينكار را تكرار كرده تا موقعي كه ديگر DNA روي كاغذ واتمن مشاهده نشود ( با نور UV آزمايش شود )
11- اين بافر را جمع آوري كرده و با P.C.I استخراج كرده و با الكل تغليظ كنيد
ب) احياي DNA از روي ژل آگارزLow Melting Point (LMP)
1- DNA مورد نظر را با آگارز LMP 2درصد الكترو فورز كنيد و اجازه دهيد تا نوارهاي DNA خوب تفكيك شوند
2- با نور UV ( طول موج 254 نانومتر موجب اسيب DNA ميشود و بهتر است از طول موج 312 نانومتر استفاده گردد ) band مورد نظر را مشاهده كنيد
3- band مورد نظر را با تيغ اسكالپر تميز علامت گذاري كنيد
4- ژل را روي كاغذ سفيد قرار داده و band مورد نظر را بريده به لوله تميرديگر منتقل كنيد
5- قطعه ژل را يك شب در فريزر 20- درجه قرار دهيد
6- قطعه ژل را در حرارت اتاق قرار دهيد و سپس 5 دقيقه در آب 65 درجه انكوبه كنيد
7-آن را به حرارت اتاق منتقل كرده و دو مرتبه با P.C.I استخراج كنيد تا ژل از آن حذف شود
8.- استات سديم ( غلظت نهايي 0.3M ) اضافه كرده با الكل رسوب دهيد
كلونينگ ( Cloning )
17- اتصال دو DNA به يكديگر در آزمايشگاه :
براي اتصال دو DNA به يكديگر در آزمايشگاه از آنزيم DNA ligase استفاده ميشود DNA ligase آنزيمي است كه بين دو نوكلئوتيد اتصال فسفودي استر ايجاد ميكند. دو نوع DNA ligase وجود دارد يكي T4 DNA ligase كه از فاژ T4 استخراج ميشود وانرژي لازم براي فعاليت خود را از ATP تامين ميكند اين آنزيم دوانتهاي blunt وهمچنين دوانتهاي Cohesive را بهم متصل كند( اتصال فسفودي استر ايجاد ميكند). ديگري DNA ligase باكتري E.coli كه انرژي لازم براي فعاليت خودراازNAD بدست مي آورد و بيشتر براي اتصال انتهاهاي Cohesive استفاده ميشود.
عمل Ligation طي سه مرحله انجام ميگيرد :
در مرحله اول گروه آدنيليل از ATP ) NAD ) به آنزيم متصل ميشود و زنجيره جانبي NH2 اسيد آمينه ليزين آدنيله ميشودو پيرو فسفات (با آنزيم T4 DNA ligase ) يا نيكو تيناميد (با آنزيم E.coli DNA ligase ) آزاد ميشود.در مرحله دوم گروه آدنيليل به انتهاي 5`-P متصل ميشود . در مرحله سوم اتصال فسفو دي استر بين گروه هيدروكسيل 3`- OH و گروه فسفوريل آدنيله شده 5` برقرار ميشودوAMP آزاد ميگردد.
منبع: http://www.iranbiotech.com
/س
در کلونینگ ژن برای انتقال ژن مورد نظر به سلول میزبان معمولاً لازم است که آن را وارد یک ناقل یا وکتور DNA ای کنند.
برای آنکه یک مولکول DNA به عنوان وکتور قابل استفاده باشد باید دارای مجموعهای از خصوصیت باشد، شامل:
1. همانندسازی مستقل از سلول میزبان داشته باشد. به این دلیل که بتواند سریعتر از تقسیم سلولی تکثیر شود تا در نهایت نسخههای زیادی از آن تولید شود.
2. در هنگام تقسیم سلولی به سلولهای دختری ( سلولهای حاصل از تقسیم ) انتقال یابد.
3. اندازهی کوچک در حدود 10 کیلوباز داشته باشد، چراکه مولکولهای بزرگتر از این مقدار ممکن است در طی مراحل تخلیص و کلونینگ خرد شود و دستورزی آن نیز مشکل است.
از معدود مولکولهای DNA ای که این ویژگیها را دارا میباشند پلاسمیدها اند.باکتریها علاوه بر کرومزومشان دارای مولکولهای DNA دو رشتهای، حلقوی و کوچکی هستند که قادر به همانندسازی مستقل از کرومزوم باکتری میباشند. پلاسمیدها معمولاً حاوی 1 یا چند ژن میباشند که سبب ایجاد خصوصیات ویژهای در سلول میزبانشان میشوند. پلاسمیدها را براساس نوع ژنشان گروهبندی کردهاند:
1. پلاسمیدهای بارور که به آنها F پلاسمید میگویند این پلاسمیدها حاوی ژنی اند به نام ژن tra که سبب انتقال اطلاعات ژنتیکی از طریق همیوغی به باکتری دیگر میشوند.
2. پلاسمیدهای مقاومت یا R پلاسمیدها که حاوی ژنهای مقاومت به یک یا چند آنتی بیوتیک اند این پلاسمیدها در میکروبیولوژی بالینی مورد توجهاند چراکه فراوانی آنها در جمعیت باکتریها، در انتخاب آنتی بیوتیک و درمان بیماریهای عفونی اثر دارد.
3. پلاسمیدهای کشنده یا کولیسن سبب تولید پروتئینهایی میشود که برای باکتریهای دیگر کشندهاند.
4. پلاسمیدهای بیماریزا مانند پلاسمید Ti در ارگوباکتریوم تومفسیانس که باعث گال در گیاهان دولپهای میشود.
5. پلاسمیدهای تجزیه کننده که امکان تجزیهی مولکولهای غیر طبیعی نظیر تولوئن را به باکتری میدهند.
در کلون ژن، از خصوصیت مقاومت به آنتیبیوتیک به عنوان یک نشانگر انتخابی استفاده میکنند. نشانگر انتخابی در گزینش کلونیهایی که DNA نوترکیب را دریافت کردهاند مورد استفاده قرار میگیرند.
باکتریوفاژها دستهای دیگر از مولکولهای DNA ای میباشند که به عنوان وکتور کلونینگ مورد استفاده قرار میگیرند. باکتریوفاژها ویروسهایی اند که باکتریها را به طور اختصاصی آلوده میکنند.فاژها مانند سایر ویروسهای از یک بخش وراثتی، که شامل مولکول داکسی ریبونوکلئیک اسید(DNA) و یا ریبونوکلئیک اسید(RNA) است، و یک پوشش محافظتی پروتئینی به نام کپسید ساخته شده. مادهی وراثتی فاژها در برگیرندهی ژنهایی است که آنزیمهای دخیل در تکثیر فاژ و پروتئینهای سازندهی کپسید را کد میکنند.
به طور کلی، برای آلوده سازی باکتری، ذرهی فاژی به دیوارهی باکتری میچسبد و مادهی ژنتیکیاش را به درون سلول باکتری تزریق میکند. سپس مولکول DNA فاژی همانندسازی میکند و بعد از ساخت اجزای سازندهی باکتری، ذرات فاژی جدید تشکیل میشود و سرانجام سلول لیز میشود که سلول را لیز میکنند و آزاد میشوند.
فاژ لاندا
همان طور که در قسمت اول گفته شد از باکتریوفاژها نیز میتوان به عنوان ناقل DNA هدف ژن کلونینگ استفاده کرد. از جمله فاژهای مورد استفاده در مهندسی ژنتیک، فاژ لاندا است که باکتری اشرشیاکولی را آلوده میکند.
وکتورهایی که از باکتریوفاژ لاندا ساخته میشوند برای کلون کردن قطعات DNA بزرگتر از 25 کیلوباز مناسبتر از وکتورهای پلاسمیدی اند. از این رو استفاده از وکتورهای مشتق از فاژ لاندا برای ساخت کتابخانهی ژنومی ( مجموعهی جامعی از قطعات DNA یا mRNA رونویسی شده از روی آنها، که نمایندهی ژنوم موجودات است) متداولتر است.
نحوهی آلودهسازی فاژ لاندا به دو صورت میتواند باشد:
1. چرخهی لیتیک- در این نوع رشد DNA فاژ در سلول باکتری همانندسازی میکند وبلافاصله سنتز پروتئینهای ساختمانی ( پروتئینهای تشکیل دهندهی کپسید یا پوشش ویروسی) از روی آن آغاز میشود و اجزای تشکیل دهندهی فاژ ( پروتئینهای کپسید و DNA فاژی ) با هم جمع شده و تشکیل یک فاژ کامل جدید را میدهند. ذرات فاژی جدید با تخریب سلول باکتری آزاد میشوند.
2. چرخهی لیزوژنیک- برخلاف چرخهی لیتیک، که DNA فاژ در سلول باکتری باقی نمیماند، در چرخهی لیزوژنیک DNA فاژی به درون کرومزوم باکتری وارد میشود در این حالت به آن پروفاژ میگویند که شکل غیرفعال فاژ است. باکتری که به این صورت آلوده شده است را لیزوژن میگویند که از نظر ظاهری تفاوتی با باکتریهای آلوده نشده ندارد. در حالت لیزوژنیک، فاژ باکتری را تخریب نمیکند و باکتری قادر به ادامهی حیات خود است. زمانی که پروفاژ از کرومزوم باکتری خارج شود فعال میشود و چرخهی لیتیک را آغاز میکند که در نهایت سبب تخریب سلول باکتری میشود. در شکل زیر نحوهی آلوده ساز فاژ لاندا نشان داده شده:فاژ لاندا شامل یک سرچندوجهی -که DNA فاژ در آن واقع شده- و یک دم -که با اتصال به دیوارهی باکتری در تزریق DNA فاژ به باکتری نقش دارد- میباشد. لانداDNA چهل و نه کیلوبازی و دورشتهای است و یک نسخه از آن به صورت خطی در قطعهی سر فاژ قرار گرفته.
نکتهی جالب در مورد ژنوم لاندا وجود دو انتهای چسبنده مکمل در طرفین مولکوی خطی DNA است. این دو انتهای چسبنده میتوانند با یکدیگر تشکیل پیوند دهند و مولکول به شکل حلقوی مبدل شود.
ژنوم لاندا پروتئینهای سازندهی سر و دم، پروتئینهای دخیل در همانندسازی و پروتئینهای فعال کنندهی چرخهی لیتیک را کد میکند. یکی از خصوصیات نقشهی ژنتیکی لاندا این است که ژنهایی که در فعالیتهای مشابه شرکت دارند در کنار یکدیگر واقع شدهاند. میتوان 4 منطقه روی آن تعریف کرد که به ترتیب قرارگیری عبارتند از: منطقهی سری ( ژنهای پروتئینهای سازندهی سر)، منطقهی دمی ( ژنهای پروتئینهای سازندهی پروتئینهای دمی) منطقهی همانندسازی، منطقهی لیتیک.
ناحیهی مرکزی ژنوم لاندا حاوی ژنهای غیر ضروری در روند آلودهسازی سلول باکتری است که آن را خارج مینمایند و قطعهی DNA خارجی را به جای آن وارد میکنند. این ناحیهی غیرضروری شامل ژنهایی است که در وارد شدن فاژ به کرومزوم باکتری و خارج شدن پروفاژ از ژنوم باکتری فعالیت میکنند.
حذف این ناحیه از دو جهت سودمند است:
1. اندازهی فاژ کوچک میشود و امکان اضافه نمودن DNA جدید با اندازهی 25 کیلوباز را به ما میدهد.
2.حذف این ناحیه فاژ را غیر لیزوژنیک کرده و باعث میشود فاژ چرخهی آلودهسازی لیتیک را دنبال کند و دیگر نیازی به القای فاژ برای تبدیل فاژ لیزوژنیک به لیتیک نیست.
بعد از وارد کردن قطعه DNA خارجی به ژنوم فاژ، آن را در پوشش فاژ بسته بندی میکنند و در مجاور باکتریهای اشرشیاکولی قرار میدهند تا آلودهسازی صورت گیرد. باتخریب سلول باکتری پلاکهایی در محیط کشت حاصل میشود به این ترتیب کلونیهای نوترکیب قابل شناسایی اند و دیگر نیاز به استفاده از نشانگر انتخابی، که در هنگام استفاده از پلاسمیدها لازم بود، نیست.
برای آنکه ژنوم نوترکیب لاندا در ساختمان سر فاژ قابل بسته بندی باشد کل اندازهی آن نباید بیشتر از 52 کیلوباز باشد. در غیر اینصورت بسته بندی انجام نشده و ذرات فاژی آلوده کننده تشکیل نمیشوند.
برای رفع نواقص این نوع وکتور و کارایی بیشتر، مهندسین ژنتیک با ایجاد تغییراتی وکتورهای مختلفی از آن ساخته اند.
فاژ M13
M13 یک باکتریوفاژ رشتهای است که متشکل است از DNA تک رشتهای حلقوی با طول 6.4 (6404 نوکلئوتید) که با کپسیدی حاوی حدود 2700 نسخه پروتئین P8 (پروتئین اصلی تشکیل دهندهی کپسید) و 5 تا 8 نسخه از پروتئین P3 که در بخش تحتانی ساختمان فاژ واقع شده است.M13 باکتری اشرشیاکولی را آلوده میکند. نحوهی ورود M13 به سلول میزبان از طریق پیلی (pili) است. پروتئین P3 به گیرندههای واقع در نوک پیلوس1 متصل شده و سبب تزریق DNA ویروس به میزبان میشود.
چرخهی آلوده سازی M13 غیر لیتیک2 است و از این رو عفونت با آن برای میزبان کشنده نیست. از طرفی DNA تک رشتهای M13 وارد کرومزوم باکتری نمیشود و چرخهی لیزوژنیک را دنبال نمیکند.DNA تک رشتهای حلقوی M13 حاوی 10 ژن است برخی از این ژنها به شرح زیر است:
• ژن II آنزیم نیکاز، که در همانندسازی چرخهی غلتان نقش دارد را کد میکنند.
• ژن III پروتئین P3 را کد میکنند که ورود ژنوم فاژ را به سلول میزبان، رهبری میکنند.
• ژن VIII که P8 را کد میکند که سازندهی اصلی پوشش ویروس است.
M13 در طی چرخهی آلوده سازی خود دو نوع روش همانندسازی را ممکن است در پیش بگیرد:
1. مولکول تک رشتهای به عنوان الگو برای سنتز رشتهی مکملش قرار میگیرد و DNA دو رشتهای حلقوی تشکیل میشود که فرم تکثیری ژنوم M13 میباشد.
2. ذرات فاژی جدید به طور مداوم تشکیل می شوند و از سلول خارج میشوند. برای آنکه ذرات فاژی جدید تشکیل شود از طرق همانندسازی چرخهی غلتان، DNA حلقوی تک رشتهای ایجاد میشود.
در همانندسازی چرخهی غلتان، فرم تکثیری ( فرم دو رشتهای حلقوی ) از طریق آنزیم نیکاز،که توسط ژن II کد میشود، در یکی از رشتهها شکاف ایجاد میشود که سبب فراهم شدن دو انتهای آزاد 3" هیدروکسیل و 5" فسفات میشود. از آنجا که آنزیم DNA پلیمراز برای ردیف کردن نوکلئوتیدها به انتهای آزاد 3" هیدروکسیل به عنوان سوبسترا نیاز دارد، عمل آنزیم نیکاز این سوبسترا را برای آنزیم فراهم میکند. رشتهی بریده نشده به عنوان الگوی همانندسازی خواهد بود. در شکل زیر دو نوع همانندسازی نشان داده شده است:
اگر M13 را با فاژ لاندا مقایسه کنید متوجهی ویژگیهای منحصربه فرد آن میشوید برای مثال داشتن DNA حلقوی تک رشتهای که طول آن خیلی کوچکتر از ژنوم لانداست و این بدین معنی است که تعداد ژنهای M13 کمتر است، به این علت که تعداد پروتئینهای سازندهی پوشش M13 تنها 3 نوع است در حالیکه در ساختمان سری دمی لاندا 15 نوع پروتئین وجود دارد.از جمله خصوصیاتی که M13را مورد توجه مهندسان ژنتیک و طراحان وکتور قرارداده است به قرار زیر است:
• اولین خصوصیت، اندازهی ژنوم M13 است. طول کمتر از 10 کیلوباز اندازهی بسیار مطلوبی برای یک وکتور است، که در قسمت قبل این مجموعه علت آن بیان شد.
• ویژگی منحصر به فرد M13، فرم تکثیری دو رشتهای و نحوهی همانندسازی آن است. فرم تکثیری M13 شبیه پلاسمید است و میتوان مانندپلاسمید، وکتور حاصل از آن را از سلول میزبانش خالص نمود و مجدداً به باکتری وارد نمود.
یک ویژگی مهم و استثنایی وکتورهای حاصل از M13 توان بالقوهی آن برای ایجاد DNA تک رشتهای از طریق همانندسازی چرخهی غلتان است. تولید ژن کلون شدهی تک رشتهای برای برخی از اهداف از قبیل توالی یابی، ایجاد جهش به منظور بررسیهای تکاملی و یا تولید پروتئینهای با ویژگیهای برتر، مورد توجه است.
1. پیلوس ساختار است که باکتری از طریق آن همیوغی جنسی انجام میدهد. پلاسمید F که حامل فاکتور بارور سازی است کد کنندهی پروتئینهای لازم برای پیلوس همیوغ کنندهی اشرشیاکولی است.
2. چرخهی لیتیک سبب لیز شدن سلول میزبان و مرگ آن میشود.
کلونینگ ژن ترنسفورماسیون
بعد از تهیه و طراحی ناقل مناسب و اتصال ژن مورد نظر به آن، DNA نوترکیب باید وارد سلول زنده شود تا با استفاده از دستگاه همانندسازی موجود در سلول، ژن هدف تکثیر و بیان شود.
عمل انتقال DNA نوترکیب به سلول زنده «ترنسفورماسیون» است و موجودی که از این طرق DNA نوترکیب را دریافت میکند تراریخت یا ترنسژنیک(Transgenic) نام دارد.سلولهایی که معمولاً به عنوان پذیرندهی مولکول نوترکیب مورد استفاده قرار میگیرند، باکتریها هستند چراکه رشد و تکثیر در باکتریها به سرعت صورت میگیرد و کشت آنها نیز سادهتر و ارزانتر است.
اکثر گونههای باکتریایی توانایی جذب مولکول DNA از محیط اطرافشان را دارند. DNA خارجی در باکتری اغلب تجزیه میشود اما در صورت داشتن جایگاه شروع همانندسازی قابل شناسایی توسط سیستم همانندسازی باکتری، میتواند در سلول میزبان باقی بماند و تکثیر شود.
قدرت جذب DNA خارجی برای همهی باکتریها به یک میزان نمیباشد. به عنوان مثال باکتری اشرشیاکولی، که پرکاربردترین باکتری در مهندسی ژنتیک و بیوتکنولوژی میکروبی است، در شرایط عادی قدرت جذب کمی دارد و میزان وقوع تراریختی در این شرایط مطلوب نخواهد بود. از این رو برای بهینه سازی میزان تراریختی باید باکتری را تحت تیمارهای خاصی قرار داد تا قدرت جذبش افزایش یابد.
در دههی 1970 با مشاهدهی اینکه سلولهای ای.کولای خیسانده شده در محلول نمکی بسیار سرد، نسبت به سلولهایی که با محلول نمکی تیمار نشده اند، DNA را خیلی بهتر جذب میکنند، راه حل مشکل موجود در مرحلهی ترنسفورماسیون یافت شد. به این ترتیب که باکتریها را در محلول کلرید کلسیم یا کلرید ربیدیوم با غلظت 50 میکرومولار قرار میدهند و سپس DNA نوترکیب را به باکتری تیمار شده میافزایند.
کلرید کلسیم تنها سبب اتصال DNA به دیوارهی باکتری میشود و به طور مستقیم در جذب DNA نقش ندارد. برای جذب واقعی DNA به درون سیتوپلاسم باکتری، آن را در معرض شوک حرارتی قرار میدهند که سبب افزایش نفوذپذیری دیوارهی باکتری میشود. حرکت DNA به درون باکتری با بالا بردن دمای انکوباتور تا 42°C تسریع میشود.
گام بعدی در کلونینگ ژن، شناسایی تراریختها است. از آنجا که در مرحلهی تولید DNA نوترکیب، تقریباً کنترلی در نحوهی اتصال قطعهی DNA به مولکول ناقل وجود ندارد، در پایان مرحله مخلوطی از انواع اتصالها، از جمله؛ مولکول ناقل فاقد قطعهی DNA ، قطعات DNA هدف متصل نشده به ناقل و مولکولهای ناقلی که به صورت نامطلوب تولید شده اند، حاصل خواهد شد. همچنین در مرحلهی ترنسفرماسیون برخی سلولها DNA نوترکیب را دریافت نمیکنند و کلونی حاصل از رشد آنها ژن مورد نظر را دربر ندارد. از این رو لازم است تا کلونیهایی که DNA نوترکیب مطلوب را دریافت کردهاند به طریقی شناسایی شوند. برای این منظور مهندسان ژنتیک، تکنیکهای مختلف طراحی وکتورها یا ناقلهای مورد استفاده در کلونینگ را متناسب با هدفی که در کلونینگ ژن دنبال میکنند، مورد استفاده قرار میدهند که در بخش دیگری به توضیح آن میپردازیم.کلونینگ DNA کاربردهای متعددی دارد که در مجموع به آنها «مهندسی ژنتیک» گفته میشود که از آن جمله تعیین توالی ژنوم موجودات است که راه را برای تعیین توالی پروتئینهای موجودات و استفاده از آن در مطالعهی عملکرد پروتئینها و مهندسی پروتئین میگشاید.
همچنین در تهیهی «اثر انگشت DNA » برای تشخیص هویت، تشخیص بیماریهای وراثتی، ژن درمانی، تولید پروتئینهای نوترکیب، ایجاد موجودات تراریخت، تولید انبوه تجاری پروتئینهای مختلف از جمله انسولین انسانی، هورمون رشد و آنزیمهای مورد استفاده در صنعت کاربرد دارد.
ناقلهاي ژن کلونينگ براي يوکاريوتها
پلاسميد 2 ميکرومتري
در قسمتهاي قبل 3 نوع از حاملين يا ناقلهاي ژني مورد استفاده در باکتري اشرشياکولي که شامل پلاسميد باکترياي، فاژ لاندا و فاژ M13 بود را معرفي کرديم.
در بيشتر مطالعات ژنتيکي و زيست مولکولي مثل مطالعهي ساختار و فعاليت ژنها از اي.کولاي که يک پروکاريوتي است به عنوان ميزبان استفاده ميشود به طوري که حاملهاي ژني متنوعي براي آن طراحي و ساخت شده است.اما براي برخي از اهداف بيوتکنولوژي، نظير خلق GMOها، استفاده از يک ميزبان يوکاريوت ضرورت مييابد. با توجه به تفاوتهاي دستگاههاي بيان ژن و ويرايش RNA که در سلولهاي پروکاريوتي ( شامل تمام باکتريها) و يوکاريوتي(شامل سلولهاي گياهي، جانوري و انواع آغازيان) وجود دارد بنابراين لازم است حاملهاي ژني ديگري را به کار برد.
مخمر ساکرومايسس سروزيه، که مخمر مورد استفاده در تخمير و نانوايي است، يکي از مهمترين موجودات در بيوتکنولوژي ميباشد.
ساکاورومايسس سروزيه گونهاي مخمر جوانهاي است که داراي تکثير غير جنسي، به طريق جوانه زدن است. اين مخمر از گذشتهي دور مفيدترين مخمر براي بشر بوده است چرا که در تخمير خمير نان و الکل سازي نقش اساسي دارد.امروزه اين تک ياختهي يوکاريوتي علاوه بر کاربرد سنتي خود به عنوان يک ميزبان يوکاريوتي براي توليد محصولات دارويي نوترکيب و پروتئينهاي يوکاريوتي از ژنهاي کلون شده اهميت ويژهاي يافته است. لذا تلاشها زيادي براي ساخت حاملين ژني براي اين مخمر صورت گرفته است.
با کشف پلاسميدي به نام پلاسميد 2µm در مخمر، که از معدود پلاسميدهاي يافت شده در سلولهاي يوکاريوت است، پيشرفت در ساخت انواع ناقلين کلونينگ براي مخمر، تسريع شد.
پلاسميد 2µm که در اکثر سويههاي ساکرومايسس سروزيه يافت ميشود و 70 تا 200 نسخه از آن در سلول مخمر موجود ميباشد در واقع خود اساس و منشا ساخت انواع ناقلهاي کلونينگ يوکاريوتي است.
پلاسميد 2µm به اندازهي 6 کيلوباز ميباشد که اين اندازه براي يک حامل ژني بسيار مطلوب ميباشد. همانندسازي پلاسميد 2µm، برخلاف همانندسازي پلاسميدهاي باکتريايي، کاملاً تحت کنترل سيکل تقسيم سلولي است و دستگاه پروئيني که اين پلاسميد و کرومزومهاي مخمر را همانندسازي ميکند يکسان است و پروتئينهاي آن توسط ژنهاي هستهاي کد ميشوند.
پلاسميد 2µm به طور کامل تعيين توالي و ژنهاي آن مشخص شده است. اين پلاسميد داراي يک جايگاه شروع همانندسازي، و 4 ژن به نامها REP1, REP2, FLP1 و D است. پروتئين حاصل از REP1, REP2 در تکثير پلاسميد دخالت دارند و پروتئين کد شده توسط ژن FLP1 با ايجاد نوترکيبي درون مولکولي ترتيب ژني را بازآرايي ميکند.
با وجودي که اندازهي کوچک اين پلاسميد و همچنين فراوانيش در سلولهاي مخمر، آن را گزينهي مناسبي براي کاربردهاي مهندسي ژنتيک کرده است اما استفاده از آن چندان هم بي دردسر نيست. از جمله مشکلات پيش رو در کلونينگ ژن به وسيلهي پلاسميد 2µm حل مسئلهي نشانگر انتخابي است. چراکه وجود نشانگر انتخابي براي جداسازي ياختههاي تراريخت از آنهايي که DNA نوترکيب را دريافت نکرده اند ضروري ميباشد.
معمولاً براي حل مشکل گزينش تراريختها از ميزبانهايي که در اثر جهش ژنتيکي توان ساخت يکي از آنزيمهاي دخيل در مسير متابوليسمي اسيدآمينهاي را از دست داده اند استفاده ميکنند. اين ميزبان را در اصطلاح «جهش يافتهي اگزوتروف» ميگويند چرا که براي ادامهي حياتش لازم است که اسيدآمينههايي که در مسير متابوليسمي آنها نقص ايجاد شده است به محيط کِشتش اضافه شود.
با وارد کردن ژن سالم آنزيمي که در ميزبان جهش يافته است به ناقل ژن ميتوان تراريختها را غربال کرد. به اين ترتيب که با کشت سلولها در «محيط کشت حداقل» (محيط فاقد هرگونه اسيدآمينه) تنها تراريختها، که از طريق DNA نوترکيب ژن سالم را دريافت کرده اند، قادر به ادامهي حيات هستند و غير تراريختها نابود ميشوند.
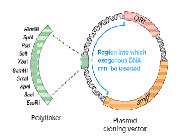
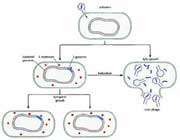



در يونان باستان اجراي هنر نمايش يكي از سرگرمي هاي خاص بود و البته يكي از اين شخصيت هاي نمايش هاي كمدي كه نماد مهمان ناخوانده را ايفا مي كرد جناب انگل بود كه نقش فردي مزاحم و آويزان را ايفا مي نمود كه بعد ها اين اصطلاح جهت تعريف نوع خاصي از روش زيست جانداران به كار برده شد.
تصوير فوق از كتاب :
تاريخچه گزارشات و كنترل بيماري هاي حيواني قابل انتقال به انسان (با بررسي و رويكرد تاريخي) اثر ژان بلانكو مي باشد كه تصويري از يك ماسك سفالي كشف شده از شهر ميرينا در يونان است كه در سال ۱۰۰ قبل از ميلاد ساخته شده است و جهت نماياندن شخصيت انگل به كار برده مي شده است .

روشهای الکتروفورز
پس ا ز استخراج مادة ژنتیک، مرحله بعدی تفکیک آن به قطعات تشکیل دهنده و شناسایی هر قطعه می باشد. همچنین در هنگام بررسی پروتئینهای سلولی نیز نیاز به تفکیک انواع پروتئینها از هم می باشد. به سبب اینکه ماکرومولکولهای زیستی باردار هستند می توان با قرار دادن آنها در یک میدان الکتریکی ، آنها را بر اساس خواص فیزیکی مانند شکل فضایی ، وزن مولکولی و بار الکتریکی ، تفکیک کرد. برای این منظور از روشی بنام الکتروفورز استفاده می شود . روشهای مختلف الکتروفورزی برای تفکیک و مطالعه بیومولکول ها اعم از اسید های نوکلئیک یا پروتئین ها ابداع شده است که در زیر به معرفی انواع متداول آن می پردازیم.
1)الکتروفورز سطحی : از یک نوار کاغذی یا استات سلولزی بعنوان فاز ثابت استفاده می شود. دو روش فوق جز در برخی کارهای پژوهشی کاربرد چندانی ندارند.
۲)الکتروفورز ژل : از یک محیط نیمه جامد (ژل ) بعنوان فاز ثابت استفاده میشود. این نوع الکتروفورز برحسب نوع ژل به کار گرفته شده به دو نوع الکتروفورز ژل پلی اکریل آمید ( PAGE) و الکتروفورز ژل آگارز تقسیم می شود. الکتروفورز PAGE دارای قدرت تفکیک بسیار بالائی بوده و برای تفکیک پروتئین ها و اسید های نوکلئیک به کار گرفته می شود.
به منظور بررسی پروتئین ها با استفاده از PAGE ، به سبب اینکه پروتئینها دارای بار مختلف هستند، معمولاً برای اینکه تفکیک فقط براساس وزن مولکولی انجام شود به بافر ماده شیمیائی SDS (سدیم دو دسیل سولفات ) اضافه می شود. SDS مولکول بزرگی با بار منفی می باشد. این ماده باعث واسرشت شدن پروتئینها شده و به آنها متصل می شود. به ازای هر دو اسید آمینه، یک مولکول SDS به پروتئین متصل می شود که باعث القاء بارمنفی متناسب با وزن مولکولی به پروتئین می شود. هر چه غلظت پلی اکریل آمید بیشتر باشد قدرت تفکیک ژل بیشتر خواهد بود و مولکول های دارای وزن مولکولی نزدیک به هم را بهتر تفکیک می نماید .
برای تفکیک اسیدهای نوکلئیک در صورت امکان از ژل آگارز استفاده می شود. تهیه ژل مزبور به مراتب سریعتر و اسانتر از ژل پلی اکریل آمید بوده و هزینه کمتری را در بر می گیرد. معمولا برای تفکیک قطعات بزرگ DNA (بزرگتر از 500 جفت باز) در صورتیکه هدف صرفا بررسی کیفی و تفکیک باشد استفاده از ژل آگارز انتخاب اول است. برای تفکیک قطعات کوچک DNA دو رشته ای و قطعات DNA تک رشته ای از ژل پلی اکریل آمید استفاده می شود. قدرت تفکیک ژل های مزبور ارتباط مستقیمی با غلظت آنها دارد. برای مثال، برای تفکیک قطعاتی به اندازه 100 جفت باز از آگاروز 3% و برای قطعات حدود 2000 جفت باز از آگارز 8/. درصد استفاده می شود.در صورتیکه نیاز به تفکیک DNA به صورت تک رشته ای باشد، از مواد واسرشت کننده نظیر اوره، فرمالدهید یا فرمامید در ژل همزمان با الکتروفورز استفاده می شود. به این نوع ژلها، ژل واسرشت کننده می گویند. چنین ژل هائی پیچ و تابهای اسید های نوکلئیک را از هم باز کرده و بنابراین تفکیک مولکول ها فقط براساس طول و نه ساختار دوم انجام می شود. در این ژل ها مولکولهای کوچکتر در مقایسه با مولکول های بزرگتر سریعتر حرکت کرده و مسافت بیشتری را طی می کنند. از روش PAGE برای بررسی موتاسیون ها و تعیین توالی DNA استفاده می شود.
۳)الکتروفورز در میدان الکتریکی ضربان دار: چنانچه اشاره شد، هر چه غلظت ژل کمتر باشد با آن می توان مولکولهای بزرگتری را بوسیلة الکتروفورز ، تفکیک کرد. ولی در این رقت محدودیت وجود دارد به طوریکه حتی با رقیقترین ژلهای آگاروز هم نمی توان مولکولهای DNA بزرگتر از 100 کیلو جفت باز را تفکیک کرد. برای این منظور از روشهای الکتروفورز در میدان الکتریکی ضربان دار استفاده می شودکه دارای انواع مختلفی است:
آ)الکتروفورز در میدان الکتریکی ضربان دار تک جهتی (UPFGE ): یکی از ساده ترین روشهای الکتروفورز در میدان الکتریکی ضربان دار، روش UPFGE می باشد. برای این روش از یک دستگاه ژل الکتروفورز افقی معمولی استفاده می شود. در UPFGE ، میدان الکتریکی در فواصل زمانی معین قطع شده و دوباره برقرار می شود. هنگامی که میدان برقرار است مولکول DNA بصورت گسترده در آمده و در جهت میدان حرکت می کند. با قطع شدن جریان الکتریکی، حرکت DNA متوقف شده و DNA فرصت می یابد تا به حالت هیئت فضایی خاص خود در آید ولی با برقراری مجدد جریان حرکت مولکول آغاز می شود. در زمانی که میدان الکتریکی برقرار نیست همة مولکولها فرصت نمی کنند بطور کامل به شکل فضایی خود در آیند هر چه مولکول کوچکتر باشد سریعتر می تواند به شکل فضایی خود در آید و از آن خارج شود. براین اساس می توان مولکولهای نسبتاً بزرگ را از هم تفکیک کرد. با استفاده UPFGE می توان مولکولهایی تا اندازه 400 کیلو جفت باز را از هم تفکیک کرد. از این روش الکتروفورزی در انالیز ژن های بزرک نظیر ژن کد کننده پروتئین دیستروفین که در پیدایش بیماری دیستروفی عضلانی نقش دارد، استفاده می شود.
ب)الکتروفورز در میدان الکتریکی معکوس (FIGE) : در روش FIGE، جهت میدان الکتریکی در فواصل زمانی معین ، معکوس می شود در این روش برای تفکیک مولکولهای کوچک از دامنه های زمانی کم استفاده می شود یعنی میدان الکتریکی بطور متناوب به مدت .5/. ثانیه در جهت مستقیم و به مدت 25/. ثانیه در جهت مخالف برقرار می شود. در مورد مولکولهای بزرگتر از زمانهای بیشتر ، 3 ثانیه به جلوو 1 ثانیه در جهت مخالف استفاده می شود. با این روش می توان مولکولهایی تا اندازه 800 کیلو جفت باز را از هم تفکیک کرد.
ج)ژل اکتروفورز در میدان الکتریکی ضربان دار (PFGE:( PFGE یکی از متداولترین روشهای الکتروفورز در میدان الکتریکی ضربان دار است. در این روش از دو میدان الکتریکی که نسبت بهم بصورت عمود قرار گرفته اند، در فواصل زمانی معین بطور متناوب استفاده می شود. یکی از این میدانها در جهت بالا به پائین و دیگری در جهت چپ به راست قرار دارد. مولکولهایDNA پس از آنکه تحت تأثیر میدان اول حرکت کردند در اثر میدان دوم 90 درجه تغییر جهت داده و حرکت می کنند. در اثر این روش مولکولهای DNA با یک حرکت چرخشی خود، در طول ژل بصورت زیگ زاگ حرکت می کنند. در روش PFGE می توان مولکولهای بسیار بزرگی در اندازه بین 20 هزار تا 2 میلیون جفت باز را از از هم تفکیک کرد.
تاریخچه ی الکتروفورز دو بعدی
الکتروفورز روشی است که در آن نمونه هایی که بار الکتریکی دارند، تحت تأثیر یک میدان الکتریکی از میان شبکه ای متخلخل حرکت می کنند و از یکدیگر جدا می شوند. سیر تکوین وتکامل این روش در جداسازی پروتئینها ارتباط تنگاتنگی با تلاشهای انجام شده در بررسی پروتئینهای سرم دارد.
در سال۱۹۳۷، "Tiselius" روشی برای جداسازی الکتروفورتیک پروتئینهای سرم ابداع کرد. انگیزه اصلی برای طراحی این روش، که بعدها به الکتروفورز منطقه ای " Zone Electrophoresis" مشهور شد، مشکلاتی بود که درحین بررسی پروتئینهای سرم وجود داشت. "Tiselius" برای اولین بار فراکشن های آلفا ، بتا، و گاما را در سرم تشریح و نام گزاری کرد. وی به خاطر تحقیق بر روی الکتروفورز و آنالیز جذبی، بخصوص اکتشافاتی که ماهیت پیچیده ی پروتئینهای سرم را مشخص می کرد، جایزه ی نوبل شیمی را در سال ۱۹۴۸ از آن خود کرد.
نقطه ی عطف در توسعة الکتروفورز در دهه های ۴۰ و ۵۰ زمانی روی داد که علاوه بر الکتروفورز منطقه ای دو روش دیگر نیز ظهور کردند: ایزوالکتروفوکوسینگ ", IEFIsoelectrofocusing" و ایزوتاکوفورزیز " Isotachophoresis". بنابراین در آن زمان امکان انجام آنالیزهای جداسازی الکتروفورتیک سه گانه بر روی مخلوطهای پروتئینی یا بصورت جدا یا در ترکیب با همدیگر بوجود آمد.
اولین تجارب در جداسازی توسط الکتروفورز دوبعدی به کارهای Smithies و Poulik در سال ۱۹۵۶ باز می گردد که با بکار بردنِ ترکیبی از الکتروفورز بر روی کاغذ و بر روی ژل نشاسته پروتئین های سرم را جداو تفکیک کردند.
پیشرفتهای بعدی در فناوری الکتروفورز، نظیر استفاده از پلی آکریل آمید و استفاده از شیبهای غلظتی پلی آکریل آمید، به سرعت در جداسازی های دو بعدی به کار گرفته شد. به خصوص استفاده از"IEF" در جداسازی دو بعدی این امر را امکان پذیر ساخت که جداسازی بُعد اول بر پایه خصوصیات بار الکتریکی پروتین ها صورت پذیرد. در سال ۱۹۷۰ الکتروفورز ژل پلی آکریل آمید به همراه سدیم دو دسیل سولفات " Sodium Dodecylsulfate-Polyacrylamide Gel Electrophoresis, SDS-PAGE" توسط "Laemmli" معرفی شد. ترکیب "IEF" با "SDS-PAGE" در بُعد دوم منجر به ابداع روشی گشت که پروتئین ها را بر پایه دوعامل متفاوت یعنی بار الکتریکی و جرم مولکولی از یکدیگر جدا می ساخت. این روش برای انواع متفاوتی از نمونه ها با خواص حل پذیری متفاوت انطباق یافت؛ بدین معنی که استفاده از اوره و دترجنت های غیریونی در "IEF"، امکان محلول سازی نمونه های متفاوت را فراهم نمود. بدین ترتیب تا سال ۱۹۷۵ یک سیستم الکتروفورز دوبعدی تکامل یافت که می توانست برای آنالیز مخلوط های پروتئینی بدست آمده از کلِ یک سلول یا بافت ها بکار گرفته شود. براساس این پیشرفتها "O’Farrell" درسال ۱۹۷۵ روش الکتروفورز دوبعدی را که برای جداسازی پروتئینهای "E.coli" بهینه شده بود، گزارش کرد. او در این روش از ایزوالکتروفوکوسینگ در ژلهای پلی آکریل آمید استوانه ای که حاوی ۸% اوره و ۲% "NP40" بود، در ترکیب با"SDS-PAGE" ناپیوسته ی"Laemmli" استفاده کرد.
ترکیب جداسازی بر پایه بارالکتریکی (نقطه ایزوالکتریک یاpI) وجداسازی براساس اندازه (جرم مولکولی نسبی یا Mr ) باعث می شود که پروتئین های نمونه در پهنای ژلِ دوبعدی توزیع شوند. این شگرد اساس پیشرفت و تکامل الکتروفورز دوبعدی را در ۳۰ سال بعدی تشکیل داد به طوری که تا کنون هزاران مقاله با استفاده از آن انتشار یافته است.
|
مطالب ذیل یک سری نکات آموزنده در مورد تست های آماری است که از سایت دانشگاه ادینبورگ اقتباس کرده ام به آدرس ذیل که البته اگر به آن مراجعه کنید بهتر است چون در صفحه من یک کمی جداول به هم خورده: http://www.biology.ed.ac.uk/research/groups/jdeacon/statistics/tress9.html#Checklist: Chi-squared test for categories of data Background: The Student's t-test and Analysis of Variance are used to analyse measurement data which, in theory, are continuously variable. Between a measurement of, say, 1 mm and 2 mm there is a continuous range from 1.0001 to 1.9999 m m. But in some types of experiment we wish to record how many individuals fall into a particular category, such as blue eyes or brown eyes, motile or non-motile cells, etc. These counts, or enumeration data, are discontinuous (1, 2, 3 etc.) and must be treated differently from continuous data. Often the appropriate test is chi-squared (c2), which we use to test whether the number of individuals in different categories fit a null hypothesis (an expectation of some sort). Chi squared analysis is simple, and valuable for all sorts of things - not just Mendelian crosses! On this page we build from the simplest examples to more complex ones. When you have gone through the examples you should consult the checklist of procedures and potential pitfalls. A simple example Suppose that the ratio of male to female students in the Science Faculty is exactly 1:1, but in the Pharmacology Honours class over the past ten years there have been 80 females and 40 males. Is this a significant departure from expectation? We proceed as follows (but note that we are going to overlook a very important point that we shall deal with later). Set out a table as shown below, with the "observed" numbers and the "expected" numbers (i.e. our null hypothesis). Then subtract each "expected" value from the corresponding "observed" value (O-E) Square the "O-E" values, and divide each by the relevant "expected" value to give (O-E)2/E Add all the (O-E)2/E values and call the total "X2"
Notes: Now we must compare our X2 value with a c2 (chi squared) value in a table of c2 with n-1 degrees of freedom (where n is the number of categories, i.e. 2 in our case - males and females). We have only one degree of freedom (n-1). From the c2 table, we find a "critical value of 3.84 for p = 0.05. If our calculated value of X2 exceeds the critical value of c2 then we have a significant difference from the expectation. In fact, our calculated X2 (13.34) exceeds even the tabulated c2 value (10.83) for p = 0.001. This shows an extreme departure from expectation. It is still possible that we could have got this result by chance - a probability of less than 1 in 1000. But we could be 99.9% confident that some factor leads to a "bias" towards females entering Pharmacology Honours. [Of course, the data don't tell us why this is so - it could be self-selection or any other reason] Now repeat this analysis, but knowing that 33.5% of all students in the Science Faculty are males
Note *1: We know that the expected total must be 120 (the same as the observed total), so we can calculate the expected numbers as 66.5% and 33.5% of this total. Note *2: This total must always be zero. Note *3: Although the observed values must be whole numbers, the expected values can be (and often need to be) decimals. Now, from a c2 table we see that our data do not depart from expectation (the null hypothesis). They agree remarkably well with it and might lead us to suspect that there was some design behind this! In most cases, though, we might get intermediate X2 values, which neither agree strongly nor disagree with expectation. Then we conclude that there is no reason to reject the null hypothesis. Some important points about chi-squared Chi squared is a mathematical distribution with properties that enable us to equate our calculated X2 values to c2 values. The details need not concern us, but we must take account of some limitations so that c2 can be used validly for statistical tests. (i) Yates correction for two categories of data (one degree of freedom) When there are only two categories (e.g. male/female) or, more correctly, when there is only one degree of freedom, the c2 test should not, strictly, be used. There have been various attempts to correct this deficiency, but the simplest is to apply Yates correction to our data. To do this, we simply subtract 0.5 from each calculated value of "O-E", ignoring the sign (plus or minus). In other words, an "O-E" value of +5 becomes +4.5, and an "O-E" value of -5 becomes -4.5. To signify that we are reducing the absolute value, ignoring the sign, we use vertical lines: |O-E|-0.5. Then we continue as usual but with these new (corrected) O-E values: we calculate (with the corrected values) (O-E)2, (O-E)2/E and then sum the (O-E)2/E values to get X2. Yates correction only applies when we have two categories (one degree of freedom). We ignored this point in our first analysis of student numbers (above). So here is the table again, using Yates correction:
In this case, the observed numbers were so different from the expected 1:1 ratio that Yates correction made little difference - it only reduced the X2 value from 13.34 to 12.67. But there would be other cases where Yates correction would make the difference between acceptance or rejection of the null hypothesis. (ii) Limitations on numbers in "expected" categories Again to satisfy the mathematical assumptions underlying c2, the expected values should be relatively large. The following simple rules are applied:
What can we do if our data do not meet these criteria? We can either collect larger samples so that we satisfy the criteria, or we can combine the data for the smaller "expected" categories until their combined expected value is 5 or more, then do a c2 test on the combined data. We will see an example below. Chi squared with three or more categories Suppose that we want to test the results of a Mendelian genetic cross. We start with 2 parents of genotype AABB and aabb (where A and a represent the dominant and recessive alleles of one gene, and B and b represent the dominant and recessive alleles of another gene). We know that all the F1 generation (first generation progeny of these parents) will have genotype AaBb and that their phenotype will display both dominant alleles (e.g. in fruit flies all the F1 generation will have red eyes rather than white eyes, and normal wings rather than stubby wings). This F1 generation will produce 4 types of gamete (AB, Ab, aB and ab), and when we self-cross the F1 generation we will end up with a variety of F2 genotypes (see the table below).
All these genotypes fall into 4 phenotypes, shown by colours in the table: double dominant, single dominant A, single dominant B and double recessive. We know that in classical Mendelian genetics the expected ratio of these phenotypes is 9:3:3:1 Suppose we got observed counts as follows
[Note: *1. From our expected total 80 we can calculate our expected values for categories on the ratio 9:3:3:1.] From a c2 table with 3 df (we have four categories, so 3 df) at p = 0.05, we find that a c2 value of 7.82 is necessary to reject the null hypothesis (expectation of ratio 9:3:3:1). So our data are consistent with the expected ratio. Combining categories Look at the table above. We only just collected enough data to be able to test a 9:3:3:1 expected ratio. If we had only counted 70 (or 79) fruit flies then our lowest expected category would have been less than 1, and we could not have done the test as shown. We would break one of the "rules" for c2 - that no more than one-fifth of expected categories should be less than 5. We could still do the analysis, but only after combining the smaller categories and testing against a different expectation. Here is an illustration of this, assuming that we had used 70 fruit flies and obtained the following observed numbers of phenotypes.
One of our expected categories (ab) is less than 5 (shown in bold italics in the table). So we have combined this category with one of the others and then must analyse the results against an expected ratio of 9:3:4. The numbers in the expected categories were entered by dividing the total (70) in this ratio. Now, with 3 categories we have only 2 degrees of freedom. The rest of the analysis is done as usual, and we still have no reason to reject the null hypothesis. But it is a different null hypothesis: the expected ratio is 9:3:4 (double dominant: single dominant Ab: single dominant aB plus double recessive ab). Chi-squared: double classifications Suppose that we have a population of fungal spores which clearly fall into two size categories, large and small. We incubate these spores on agar and count the number of spores that germinate by producing a single outgrowth or multiple outgrowths. Spores counted:
Is there a significant difference in the way that large and small spores germinate? Procedure: 1. Set out a table as follows
2. Decide on the null hypothesis.
3. Calculate the expected frequencies, based on the null hypothesis.
4. Decide the number of degrees of freedom
5. Run the analysis as usual. Calculating O-E, (O-E)2 and (O-E)2/E for each category, then sum the (O-E)2/E. values to obtain X2 and test this against c2 . The following table shows some of the working. The sum of the values shown in red gives X2 of 20.23
We compare the X2 value with a tabulated c2. with one degree of freedom. Our calculated X2 exceeds the tabulated c2 value (10.83) for p = 0.001. We conclude that there is a highly significant departure from the null hypothesis - we have very strong evidence that large spores and small spores show different germination behaviour. Checklist: procedures and potential pitfalls Chi squared is a very simple test to use. The only potentially difficult things about it are:
If you follow the examples given on this page you should not have too many difficulties. Some points to watch:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

شیستوزوماها به علت شکل باریک و کشیده خود و جدا بودن ۲ جنس نر و ماده از یکدیگر با سایر ترماتودها تفاوت دارند.کرم نر ،بزرگتر ،خاکستری رنگ و دارای انتهای قدامی استوانه ای است و بدن سنگین آن دارای چین هایی است که تشکیل یک کانال ژنیکوفور طویل و شکمی را می دهد که کرم ماده تیره تر و باریک تر در طی جفت گیری در آن جای می گیرد.پوشش خارجی کرم بسته به گونه ،صاف یا دارای برجستگی است.روده به ۲ سکوم تقسیم می شود که در انتهای خلفی کرم به هم پیوسته و تشکیل لوله منفرد کوری را می دهد.تعداد بیضه ها در جنس نر و طول رحم و تعداد تخم ها ، مشخصه گونه های مختلف هستند.سیستم دفعی مرکب از سلول های شعله ای ،لوله های جمع آوری کننده و ۲ لوله طویل است که به مثانه کوچکی با یک سوراخ دفعی انتهایی ختم می شوند.

چرخه زندگی
بسته به گونه کرم روزانه از ۳۰۰ تا ۳۵۰۰ عدد تخم وارد سیاهرگ ها می شود.بعضی تخم ها توسط جریان سیاهرگی به مرکز کبد فرستاده می شوند ولی بسیاری از آن ها توسط انقباض سیاهرگ ها در محل باقی مانده و در نتیجه آنتی ژن ها و آنزیم های تراوش شده از میراسیدیم داخل تخم واکنش التهابی موضعی آغاز می شود.مولکول های چسبنده ای که توسط سلول های التهابی ایجاد می شوندو به تازگی شناخته شده اند نیز ممکن است در روند فوق شرکت داشته باشند.تداوم این واکنش منجر به پارگی دیواره سیاهرگ ها و حرکت تخم ها به سوی بافت های اطراف عروقی روده و مثانه و سرانجام ورود آنها به داخل مجاری این اعضا می شوند که در این صورت تخم ها با مدفوع یا ادرار دفع می شوند.تخم پس از تماس با آب تازه باز شده و میراسیدیم خارج شده تا زمان یافتن مناسب ، در آب شنا می کند.
میراسیدیم به منظور ادامه چرخه حیات خود باید در مدتی قریب به ۱۰ ساعت وارد بدن حلزون شود.در بدن حلزون پس از ایجاد ۲ نسل از رشد و تکثیر اسپوروسیست ،که روند پیچیده ای دارد سرکر های با دم ۲ شاخه از حلزون خارج می شوند.سرکر انگل پس از ۲۴ تا ۳۶ ساعت ،عفونت زایی خود را برای میزبان مهره دار از دست می دهد.تکثیر انگل در حلزون میزبان واسط ،شانس بقای آن را افزایش می دهد.یک میراسیدیم می تواند هزاران سرکر ایجاد کند.پوست انسان در طی استحمام ،شنا ،کار و یا شستشوی لباس در تماس با سرکر های با شنای آزاد قرار می گیرد.در این صورت سرکر به پوست چسبیده و با خشک شدن آب سطح پوست به داخل بستر عروق محیطی نقب زده و همزمان با ورود به داخل پوست ،دم خود را از دست می دهد.در صورت خورده شدن سرکر با آب ،سرکر وارد غشای مخاطی دهان و گلو می شود .انگل که در این حالت شیستوزومولا نامیده می شود،در طی چند ساعت در غشای خود دچار تغییرات ریخت شناسی و بیوشیمیایی شده و به زودی خود را با آنتی ژنها ی میزبان پوشانده و به این ترتیب از حملات ایمنی میزبان فرار می کند.شیستوزومولا از طریق خون آوران به قلب راست و ریه ها حمل می گردد و سپس با عبور از مویرگ های ریوی به داخل گردش عمومی خون و سرانجام به عروق کبدی می رود.در بخش داخل کبدی سیستم پورت،ترماتود های خونی تغذیه کرده و به سرعت رشد می کنند.قریب به ۳ هفته پس از تماس پوست با سرکر ، کرم های در حال بلوغ از جریان خون پورت به داخل سیاهرگ های مزانتر ، مثانه و لگن مهاجرت می کنند.دوره نهفته برای شیستوزوما مانسونی ،۸-۷ هفته ،شیستوزوما هماتوبیوم ۱۲-۱۰ هفته و برای شیستوزوما ژاپونیکوم ۶-۵ هفته است.کرم های بالغ ممکن است تا ۳ سال در بدن انسان زندگی کنند ولی متوسط دوره زندگی آن ها احتمالا ۱۰-۳ سال است.

**انگلهاي شيستوزوما داراي سير تكاملي پيچيده و چند مرحلهاي هستند. بطوريکه كرمهاي بالغ در عروق خوني انسان مستقر شده، تخمها از طريق ادرار يا مدفوع به محيط آبي راه پيدا ميكنند، ميراسيديوم كه اولين مرحله لاروي است از داخل تخم آزاد گشته و به حلزون ميزبان واسط نفوذ ميكند، سپس انگل به مراحل اسپروسيست و سركر، تبديل مي گردد و نهايتا سركر با دم دو شاخه از حلزون خارج ميشود واز راه نفوذ پوستي، ميزبان بعدي را آلوده ميكند. بنابراين ما در هر مرحله از زندگي انگل شاهد تغييرات مرفولوژيك، ساختاري و فيزيولوژيك هستيم و در يك جمله ميتوان گفت كه انگل بايد از خصوصيات تطابقي بالايي جهت ادامة سير تكاملي و بقاء خود برخوردار باشد.

به طور كلي خصوصيات تطابقي انگل با ميزبان واسط حلزون را در چند مرحلة مجزا ميتوانيم مورد بررسي قرار دهيم:
۱‐ مرحلة باز شدن تخم و آزاد سازي ميراسيديوم
۲‐ ميزبانيابي و نفوذ ميراسيديوم به حلزون
۳‐ ارتباط حلزون‐ مرحلة لاروي
۴‐ خروج سركر از حلزون واسط
امروزه مشخص شده است كه در باز شدن تخم ميراسيديوم، علاوه بر شرايط محيطي و عوامل كموكينزيس مانند نور، درجة حرارت، فشار CO2و غيره، ترشح برخي از آنزيمها از طرف ميراسيديوم مانند لوسين آمينو پپتيداز نقش بسيار مؤثري، دارد. مطالعات نمايانگر اين پديده هستند كه از ترشح اين آنزيم در بدن انسان ممانعت به عمل ميآيد، چرا كه غلظت NaCl موجود در جريان خون و بافتهاي بدن انسان به عنوان يك مهار كننده براي ترشح آنزيم عمل ميكنند و ترشح آنزيم فوق تنها وقتي صورت ميگيرد كه تخم در شرايط محيط آبي قرار گيرد. بنابر اين باز شدن تخم در بدن انسان صورت نمي گيرد. عملكرد اين آنزيم را به حل نمودن پوشش تخم نسبت ميدهند.
يكي از ساختارهاي مهم تطابقي انگل در اين مرحله، پاپيلا ميباشد كه حاوي مقادير زياد رشته هاي عصبي است و از اين طريق به راحتي ميتواند شرايط محيطي را درك كند. به علاوه ميراسيديوم داراي غدد نفوذي و رأسي ميباشد كه ترشحات اين غدد در نفوذ آن به بدن حلزون نقش بسيار موثري را ايفا ميكنند. به هر حال ميراسيديوم از طريق پاپيلاي رأسي به مواد محلولي كه از حلزون ترشح ميشود و به نام گليكوپروتئينهاي جاذب ميراسيديوم(Miracidium Attracting Glycoproteins) معروف هستند، پاسخ مي دهد و به سمت حلزون جذب مي شود.
مشخص شده است كه ناحية كربوهيدراتي اين گليكوپروتئينها خاصي سيگنالينگ براي ميراسيديوم داشته و نهايتًا ميراسيديوم بعد از چند بار برخورد با بدن حلزون از ناحية رأسي، اتصال خود را برقرار ميكند. اتصال ميراسيديوم به حلزون يك پديده اختصاصي بوده و به نظر ميآيد كه اين پديدة يک پديده وابسته به رسپتور و ليگاند است. ملكولهاي دخيل در اين امر هنوز كامل مشخصن شدهاند ولي به نظر ميآيد كه کمورسپتورهاي ميراسيديوم با گليكوپروتئينهاي موكوسي بدن حلزون اتصال مييابند. قابل ذكر است كه در اين مرحله ميتوان با بلوكه نمودن كمورسپتورهاي ميراسيديوم از آلودگي حلزون و ادامه سير تكاملي انگل جلوگيري كنيم و نقش مهمي را در كنترل داشته باشيم. نهايتًا ميراسيديوم با ترشحات پروتئازي كه از مهمترين اين ترشحات، سيستيئن پروتئاز است همراه با حركت متهاي خود وارد بدن حلزون شده و به اسپروسيست مادر تمايز پيدا ميكند.
مشخص شده است كه در طي ۲۴ ساعت، اپيتليوم مژه دار ميراسيديوم از بين ميرود و جاي خودش را به يك اپيتليوم سينسيشيال همراه با ميكروويلي هاي تخصص يافته ميدهد. اين تغيير تگومنتي انگل منجر به دريافت مواد غذايي به واسطة ميكروويلي سطحي تگومنت شده و از آنجائيكه اسپروسيست فاقد عضو گوارشي است، اين موضوع يك خصوصيت تطابقي اسپروسيست محسوب ميشود. به علاوه، تگومنت اسپروسيست قادر به جذب آنتيژنهاي گليكوپروتئيني بافت بدن حلزون مي باشد و به اين ترتيب اسپروسيست خود را از ايمني حلزون مخفي ميكند. در واقع اسپروسيست نيز به نحوي داراي مكانيزم فرار از سيستم ايمني همانند آنچه ما در كرمهاي بالغ در خون مشاهده ميكنيم، ميباشد. اما جالب آنكه، حلزونهاي واسط هم تمايل به بقاء انگل و ادامة سير تكاملي آن دارند چرا كه مشخصشده است كه حلزونهاي آلوده، توانايي جذب ميراسيديومهاي جديد را به خود از دست ميدهند و از اين طريق از آلودگي بيشتر و مرگشان جلوگيري ميكنند. نهايتًا سركر در غدد هضمي حلزون توليد ميشود.
*مشخص شده است كه خروج سركرهاي اكثرًا گونه هاي شيستوزوماي انساني از بدن حلزون در اوقات گرم روز اتفاق ميافتد، همين موضوع يك رفتار تطابقي سركر محسوب ميشود، چرا كه انسان نيز در همين زمان در تماس با آب قراردارد. امروزه مشخصشده است كه اين امر، به دليل بيان بالاي پروتئيني به نام ردوپسين در سركر ميباشد و در واقع خاصيت فوتوتروپيك بودن سركر نيز به علت وجود اين پروتئين است. به علاوه سركر به واسطة داشتن غددي تك سلولي درناحية رأسي خود، به راحتي ميتواند از بدن حلزون خارج شود و خود را به ميزبان اصلي برساند. تگومنت سركر نيز دستخوش تغييراتي ميشود و آن وجود پوشش به نام گليکوكاليكس است كه منجر به حفظ سركر در محيط و بقاء آن در شرايط محيط آبي ميگردد. لازم به ذكر است كه تغييرات تگومنتي در هر مرحله از سير تكاملي انگل مشاهده ميشود و اين امر يكي از مهمترين رفتارهاي تطابق انگل محسوب ميشود. تطابق انگل در ميزبان نهايي را نيز ميتوانيم در چند قسمت مجزا مورد بررسي قرار دهيم:
۱‐ جذب سركر به سطح پوست
۲‐ نفوذ سركر از لايه هاي مختلف پوست
۳‐ ترانسفورماسيون سركر به كرم جوان
۴‐ تطابق شيستوزومولوم و كرم بالغ در بدن انسان
*مشخص شده است كه ترشحات پوستي افرادي كه به آب آلوده وارد ميشوند، در جذب سركر اهميت دارند. آرژنين يكي از ترشحات پوستي است كه سبب جذب سركر به پوست ميشود. جالب اين جاست كه خود سركر هاي جذب شده به پوست نيز شروع به توليد آرژينين از غدد تك سلولي ميكنند و همين امر سبب فراخواني ساير سركرها به پوست و افزايش احتمال آلودگي فرد ميشود. از ديگر ترشحات پوستي ميتوان به
اسيدهاي چرب ضروري مثل اسيد لينولينك و اسيد لينولئيك اشاره داشت كه نقش مهمي را در جذب سركر به پوست ايفاء ميكنند. اين اسيدهاي چرب در القاء نفوذ سركر به پوست و تغييراتي كه در تگومنت بايد صورت گيرد نيز دخالت دارند.
* سركر بايد بتواند از لايه هاي مختلف پوست كه شامل لايههاي اپيدرميس، درميس و هايپودريس است، نفوذ كند و خود را به سياهرگها و عروق خوني برساند. مطالعات نشان دادهاند كه حدود ۵۰ % سركرها در اين مرحله از بين ميروند. سركر با واسطة آنزيمي به نام الاستاز كه يك نوع سرين پروتئاز است و حدود ۹۰ % از ترشحات سركر ي را به خودش اختصاصميدهد، به راحتي ميتواند از لايههاي فوق عبور كند. اين آنزيم
علاوه بر تسهيل نفوذ انگل، در حذف گليكوكاليكس نيز نقش مهمي دارد. حذف گليكوكاليكس از اهميت بالايي برخوردار بوده چرا كه اين ماده از عوامل فعال كننده سيستم كمپلمان است و انگل از اين طريق يكي از استراتژيهاي فرار از سيستم ايمني را اعمال ميكند. سركر براي عبور از لايه هاي پوست نياز به مقادير فراوان انرژي دارد و به همين دليل است كه آنزيمي به نام تريپتوفان هيدروكسيلاز به ميزان فراوان در اين مرحله يافت ميشود. اين آنزيم در تبديل تريپتوفان به سروتونين نقش مهمي دارد. سروتونين سبب افزايش دريافت گلوكز از محيط و تحريك مسير توليدانرژي در سركر ميشود و به اين ترتيب انرژي لازم براي تحرك خود را بدست ميآورد.
*مطالعات نشان دادهاند كه مواجهة سركر با دماي ºC ۳۷ ، غلظت قند و اسمولاريته منجر به ترانسفورماسيون سركر به شيستوزومولوم ميشود و به نظرميآيد كه اين سه عامل با اثر سينرژيسمي سبب القاء تحريك سركر ميشود.
*با تبديل سركر به شيستوزومولوم متابوليسم از فرم هوازي به بيهوازي تغيير مييابد و سطوح آنزيمهاي تنفسي كاهش مييابد. در اين مرحله انگل جهت پر كردن مخازن انرژي خود به جذب گلوكز با استفاده از پروتئينهاي ترانسپورتر به نام4 SGTPميپردازد. تغييرات تگومنتي نيز از مهمترين تغييرات اين مرحله محسوب ميشود. مشخص شده است كه سه ساعت بعد از نفوذ انگل لايه هاي غشايي افزايش مييابند و در طي دو هفته غشاء ضخيم شده و فرورفتگي هايي به منظور افزايش سطح جذب مواد غذايي صورت ميگيرد. البته شيستوزومولوم به حذف اكسيداتيو توسط سيستم ايمني حساس بوده و با مكانيسم ADCCاز بين ميرود. به همين دليل تعداد زيادي از كرمهاي جوان در نتيجه فعاليت سيستم ايمني از بين ميروند، مقدار كمتري نيز با به كارگيري استراتژيهاي فرار خود را به عروق خوني باب رسانيده و به سير تكاملي خود ادامه
ميدهند.به طور كلي كرمهاي بالغ و شيستوزومولومها با مكانيسمهاي زير ميتوانند خود را از دست سيستم ايمني مصون بدارند:
اتصال اجزاء سيستم ايمني مانند آنتيباديها و اجزاء كمپلمان مانند C3 كه در سطح تگومنت انگل قرار ميگيرد، به عنوان محركي جهت پرتاب لايه هاي غشايي و نو شدن آنها ميشود. در واقع سيستم ايمني ميزبان كمك به بقاء بيشتر انگل ميكند.
شيستوزما قادر به جذب آنتيباديها، اجزاء كمپلمان و گروههاي خوني در سطح تگومنت است. در اين مورد فرضيه هاي مختلف وجود دارد.امروزه مشخص شده است كه از آنجائي كه ژنوم شيستوزوما در بسياري از تواليها با ژنوم انسان تشابه دارد، انگل قادر به سنتز آنتيژنهاي ميزبان مي باشد و Ag هاي عرضه شده در سطح تگومنت همانند آنتي ژنهاي ميزبان است. به علاوه انگل توانايي سنتز اسيدهاي چرب را ندارد و از طريق پروتئيني به نام FABP (Fatty Acid Binding Protein) به LDL ميزبان اتصال مييابد و ماهيت بيگانگي خود را مخفي ميكند.
پروتئيني به نام پاراميوزين كه به عنوان رسپتور بخش IgE, FC مي باشد در سطح انگل شناسايي شده است. مطالعات نشان دادهاند كه اگر آنتي بادي مونوكلونال ضد پاراميوزين را به موش تزريق كنيم و سپس همين موش را به انگل آلوده كنيم، بلوكه شدن رسپتور FCايمونوگلوبينها در پديدة ماسك زدن انگل تداخل مييابد و باركرمي كاهش پيدا ميكند. لازم به ذکر است که اين پروتئين کانديد واکسن نيز مي باشد.
تحقيقات مويد اين مطلب هستند كه اگر عصارة انگل را بدست آوريم و در مجاورت IgE انسان يا رات قرار دهيم، اين Ab به صورت نان فانكشنال درآمده كه به دليل فعاليت الاستازي يا سرين پروتئازي انگل ميباشد. در نتيجه انگل با حذف IgE ميتواند بر سيستم ايمني غلبه كند
مطالعات نشان دادهاند كه تنها ۲% از كرمهاي بالغ در مجاورت ائوزينوفيل و IgE كشته ميشوند كه اين موضوع نمايانگر بيان بالاي آنزيمهاي آنتياكسيدان جهت زيست انگل در عروق خوني ميباشد. دو آنزيم شناخته شده در اين انگل به نامهاي سوپراكسيدديسموتاز و گلوتاتيون پراكسيداز است كه ميتواند سبب خنثي شدن راديكالهاي 2H2O2, O شود. در مطالعهاي مشخصشده است كه ميزان بيان اين آنزيمها با تكامل انگل افزايش مييابد به طوري كه بيشترين ميزان بيان آن در كرم بالغ و كمترين ميزان بيان در شيستوزومولا مشاهده شده است. جايگاه آنزيمهاياخير با استفاده از روشهاي فلورساني در غشاء اپيكال تگومنت يعني در محل برخورد سيستم ايمني با تگومنت مشخصشده است. نهايتًا انگل در هر مرحله از سير تكاملي خود با تغييرات مرفولوژيك، فيزيولوژيك ، بيوشيميايي و مولكولي سعي در ادامة تكامل خود ميكند و در واقع از اين تغييرات در جهت حفظ و بقاء خود استفاده ميكند. لازم به ذكر است كه با شناسايي ملكولهاي دخيل در اين تطابق در هر مرحله ميتوان اقدامات مهمي در جهت شناسايي داروها و طراحي واکسنها پرداخت.
PASSWORD: www.virologist.blogfa.com
References:
Ansary, A. EL., 2003, Biochemical and immunological adaptation in schistosome parasitism, comparative biochemistery and physiology part B- 136: 227- 243.
Ansary, A. Daihan, S. AL., 2005, Stage specifically expressed schistosome proteins as potential chemotherapeutic targets, Med Sci Monit, 11, 3: RA94- 103.
Tloverde, Ph., Carvalho- Queiroz, C., Cook, R, 2004, Vaccination with antioxidant enzymes consers protective immunity against challenge infection with Schistosoma mansoni, Mem Inst Oswalldo Cruz, Rio de Janeiro, 99: 37- 43
Hellemond, J., Retra, K., Brouwers, J., Balkom, B et al,2006, function of the tegument of schistosomes : clues from the proteome and lipidome, Inter J for parasitol, 36: 691- 699.
Loukas, A., Jones, M., King, L et al, 2001, Receptor for FC on the surface of schistosomes, infec and immun . 69: 3646-3651
Skelly, P., Shoemaker, Ch., 2000, Induction cues for tegument formation during the transformation of Schiostosoma mansoni cercariae, Inter J for parasitol, 30: 625- 631
Theron, A., Coustau, C., 2005, Are biomphilaria snails resistant to Schistosoma mansoni? J of helminth,79: 187- 1
دوالپا نمونه ای از اسطوره های انگلی
چند روز پیش کتاب ولنگاری صادق هدایت را می خواندم که در آن اشاره به دوالپا شده بود .موجود جالبی که کاملا از خصوصیات انگل های خارجی تبعیت می کرد.
دَوالپا یکی از موجودات خیالی در اسطورهها و داستانهای ایرانی است که بالاتنه انسان دارد و پاهایش مانند تسمه دراز و پیچندهاند. دوالپا در زبان فارسی مصداق آدمهای سمجی است که به هر دلیل به حق یا ناحق به جایی با کسی میچسبند و آنجا را رها نمیکنند چنانکه هنگامی بچهای سماجت کند و به مادر اصرار ورزد مادر به او گوید: «چرا مثل دوالپا به من چسبیدهای.»
افسانههای قدیم میگویند در بیابانهای بزرگ و سوزان، حتی در برخی جنگلهای دوردست، موجودی زندگی میکند به نام دوالپا . دوالپاها مردانی نحیف و به ظاهر درمانده بودند با پاهایی به لاغریِ تسمه (دوال) که در کنار جادهها شل و بیرمق میافتادند یا دراز میکشیدند. وقتی مسافران که در قدیم اغلب تنها و پیاده سفر میکردند به آنها برمیخوردند ناگهان میایستادند و بعد که نزدیکتر میرفتند یکه میخوردند. پیرمردی را میدیدند پاره پوش و لاغر اندام، با سر کوچک و ریش که با چشمان نیمهباز و پرالتماس به آنها نگاه میکرد: [مسافر : چه شده پدر جان ؟ در راه مانده ای ؟
(دوالپا حرفی نمیزند و فقط ناله میکند.)
مسافر: راهزنها داراییات را بردهاند؟ حیوانات درنده به تو حمله کرده اند؟ من چه کاری برایت انجام دهم؟
دوالپا: (نالان) دیگر نای راه رفتن ندارم.... پیرشی جوان، من را تا زیر سایهی آن درخت برسان؛ بعدش برو که دست خدا به همراهت! و با دستش تک درختی را که قدری جلوتر است نشان میدهد.
مسافر که میبیند پیرمرد با این همه ناتوانی چه اندک توقع است با همهی وجود مردک را از جا بلند میکند و برگردهاش سوار میکند.
دوالپا پاهای تسمه مانند چهل متری خود را که زیر بدن پنهان کرده بود گشوده گرداگرد بدن مسافر چنان میپیچد واستوار میکند. بیچاره مسافر، اول خیال میکرد به زودی و با رسیدن به سایهی درخت از فشار پاها آسوده میشود اما آنجا بود که تازه میفهمید دوالپا خیال پیاده شدن ندارد .
[ مسافر : ( نفس زنان ) رسیدیم پدرجان .... بی زحمت پیاده شو! ... گردنم خرد شد!
دوالپا : من پدرت نیستم بدبخت! من دوالپایم! کارم این است که احمق هایی مثل تو را شکار کنم و از گردهشان پایین نیایم! چرا ایستادهای؟ یالا راه بیفت ! ]
خلاصه آن که دوالپای ناجوانمرد چون بختک بر شانههای مسافر بیچاره میافتاد و به هر طرف که میخواست او را میکشاند. مسافر هم مجبور بود فرمانهای دوالپا را گردن بگذارد؛ اولین کار دوالپا این بود که مسافر را از مسیر اصلیاش خارج میکرد و در بیراهه میانداخت. دوالپا که سراسر بیابان را مثل کف دست میشناخت کوره راههای بیسرانجام و دورهای باطلی را انتخاب میکرد که راه به هیچ دهکورهای نمیبرد. بعد آذوقه و آب مسافر را تصاحب میکرد و خودش میخورد و مینوشید. و دست آخر پس از چند روز سواری پیکر نیمهجان قربانی را وسط بیابان رها میکرد تا خوراک لاشخوران شود و خودش به کنارهی راه اصلی باز میگشت و در انتظار مسافر بعدی میماند.
در روزگاران قدیم مسافر دانایی در راه گول دوالپایی را خورد و او را بر پشت خود سوار کرد . وقتی فهمید دوالپا پیش از مردن او قصد پیاده شدن ندارد، چارهای اندیشید. مسافر هندوانهای به همراه داشت که دوالپا مغزش را خورد و پوستش را به زمین انداخت. مرد پوست هندوانه را برداشت و زیر جامه پنهان کرد . چند خوشه انگور را هم که به همراه داشت دور از چشم دوالپا در پوست هندوانه انداخت. مرد در حالی که تسمهی دوالپا بر گردنش استوار شده بود چهل روز در بیابان سواری داد و زجر کشید. چهل روز تاب آورد و خودش را زنده نگه داشت؛ تا این که پس از چهل روز انگورها در پوست هندوانه تبدیل به شراب شدند! مسافر کاسهی هندوانه را از زیر جامه بیرون آورد. دوالپا پوست هندوانه را دید و قاپ زد. بعد جرعه جرعه شراب را نوشید. اندکی که گذشت دوالپا مست شد و از پشت مسافر به زمین افتاد . مرد خسته دوالپای خبیث را کشت و به سوی راه آبادی حرکت کرد ....
ازین رو دوالپا در زبان فارسی مصداق آدمهای سمجی است که به هر دلیل به حق یا ناحق به جائی میچسبند وبدون توجه به تاریخ مصرف خود آنجا را رها نمیکنند.
دَوال در فارسی به معنی تسمه است.
در کتاب سندباد بحری توصیفی اینگونه از دوالپا دیده میشود: «دوالپا پیرمردی است که دم جاده نشسته گریه میکند و هر رهگذری که میرسد به او التماس کرده میگوید مرا کول بگیر از روی نهر آب رد کن. هر کس او را کول بکند یکمرتبه سه ذرع پا مانند مار از شکمش درآمده دور آن کس میپیچد و با دستهایش محکم او را گرفته فرمان میدهد: کار بکن بده به من. برای اینکه از شر او آسوده بشوند باید او را مست کرد.»
کتاب عجائب المخلوقات و غرائب المخلوقات محمدبن محمود طوسی (قرن ششم هجری) ضمن نقل حکایتی نام دوالپا را به کار میبرد و آن را گونهای نسناس مینامد. البته در لغتنامه دهخدا، داستان وامق و عذرا را منبع دیگری هم نقل میکند. از آنجا به کتابهایی همچون سلیم جواهری و وغوغ ساهاب صادق هدایت راه یافتهاست.
بدن جانداران از سلول های متنوعی تشکیل شده است و این تنوع باعث عملکرد اختصاصی تر این سلول ها شده است. بنابراین تمام سلول ها مجبور به انجام تمام عملکردهای فیزیولوژیک یا تولید تمام انواع محصولات لازم برای بقاء فرد نیستند. هر چند اغلب این سلول ها حاوی ارگانلهای داخلی مشترکی بوده و مسیرهای متابولیک مشترکی را طی می کنند، ولی هر سلولی اجزاء یا محصولات خاصی را بطور اختصاصی تر و به مقدار خیلی زیاد تر بیان می کند . در حقیقت بیان اختصاصی این جزء، موجب عملکرد اختصاصی سلول و متمایز شدن آن از سایر سلول ها می شود. بطوریکه سلول های مختلف، توانایی های مختلفی در انقباض یا انتقال پالس های الکتریکی یا ترشح محصول خاصی نظیر هورمون ها یا آنزیمها و غیره دارند. این تخصص در عملکرد سلول ها باعث می شود که حیوانات از انواع مختلفی از سلول ها تشکیل شوند. این سلول ها از نظر خصوصیاتی نظیر اندازه، شکل، ساختمان و عملکرد سلو لی با هم متفاوت هستند. به این نوع تفاوت سلولی تمایز 1 گفته می شود. در حقیقت این سلول ها از نظر ساختار ژنتیکی یکسان بوده ولی بیان ژنهای آنها تفاوت دارد ، بطوریکه مهره داران دارای بیش از 100 نوع سلول مختلف هستند که در اعضاء مختلف با عملکردهای مختلف دیده می شوند . بسیاری از سلول های حیوانات را می توان در خارج از بدن حیوان تکثیر داد. سلول های جدا شده از بافت ها و اعضای حیوانات می توانند در دمای مناسب در یک محیط حاوی مواد غذایی و عوامل رشد درون ظروف پلاستیکی رشد کنند. کشت اعضاء، بافت ها و سلول ها ی جانداران در محیط آزمایشگاهی را کشت سلولی 2 می نامند که کاربردهای متنوعی در شاخه های مختلف علوم زیستی دارد. بسیاری از سلول های جانداران در محیط خارج از بدن قابل کشت و نگهداری هستند . به عنوان مثال می توان به سلول های بافت همبند نظیر فیبروبلاستها؛ سلول های بافت اسکلتی نظیر سلول های استخوانی و غضروفی؛ سلول های بافت عضله قلب و عضله صاف؛ سلول ها ی اپی تلیال نظیر سلول های ریه، پستان، پوست، مثانه و کلیه؛ سلول های عصبی نظیر سلول های میکروگلیال؛ سلول های اندوکرین نظیر سلول های غده آدرنال، هیپوفیز و جزایر پانکراس ؛ ملانوسیتها و بسیاری از سلول های سرطانی اشاره کرد. در حقیقت توسعه تکنیک های کشت سلولی عمدتاً مدیون تحقیقات سرطان شناسی و ویروس شناسی می باشد.
اندازه گیری حیات، رشد و تکثیر سلولها کاربردهای مختلفی در تحقیقات دارد. یکی از قدیمی ترین روشها رنگ آمیزی سلولها با تریپان بلو است که حساسیت کافی نداشته و نیاز به بررسی میکروسکوپی دارد. روشهاي ديگري نظیر اندازه گيري ميزان كل اسيدهاي هسته اي و ميزان پروتئين در ليزات سلولي نیز از دقت كافي برخوردار نمي باشند.
اندازه گیری جذب مواد رادیواکتیو توسط سلولهای در حال رشد روش دقیقی است. تركيباتی نظیر تیمیدین نشاندار شده با تریتیوم و 5- برومودئواكسي اوريدين هنگام پروليفراسيون سلول وارد ساختمان هسته سلول شده و باعث نشاندار شدن سلولها می شوند. اما این روش نیز وقت گیر بوده و بعلاوه نیازمند هاروست کردن سلولها پس از تحریک سلولی است. همچنین مشکلات مربوط به کار با مواد رادیواکتیو نظیر دفع مواد باقی مانده و حفاظت از اشعه را نیز به دنبال دارد.
در سال 1983 آزمایش MTT بعنوان جایگزینی برای روش رادیواکتیو پیشنهاد شد. در این روش آنزیماتیک بعنوان سوبستراي واكنش از نمكهاي محلول تترازوليوم كه مهمترين آنها MTT است استفاده می شود. بوسیله این آزمایش ساده و دقیق می توان پاسخ سلولهای مختلف را به فاکتورهای خارجی از جمله فاکتورهای رشد، داروهای سایتوتوکسیک و سایر عوامل شیمیائی ارزیابی کرد. بطوریکه میزان فعالیت و سرعت تکثیر سلولها ممکن است تحت تاثیر هورمونها، فاکتورهای رشد، سايتوكينها و میتوژنها افزایش یافته یا تغییری نکند. همچنین تحت تاثیر بعضی از داروها و عوامل سایتوتوکسیک نظير داروهاي ضد سرطان ممکن است سلولها دچار نکروز یا آپپوپتوز شده و سرعت تکثیر یا رشدشان کاهش یابد.
این روش نسبت به ساير روشهاي بررسي پروليفراسيون سلولي ساده تر بوده و با امكانات موجود در اغلب آزمایشگاهها قابل اجراست. بعلاوه کلیه مراحل آزمایش در پلیتهای 96 خانه کشت سلولی انجام شده و نتایج با دستگاه الیزا ریدر قرائت می شود لذا تعداد زیادی نمونه را می توان همزمان آزمایش کرد.
اساس تست MTT :
آزمایش MTT که یک روش رنگ سنجی است که بر اساس احیا شدن و شكسته شدن كريستالهاي زرد رنگ تترازوليوم با فورمول شيميائي3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide (MTT) بوسيله آنزيم سوكسينات دهيدروژناز و تشكيل کریستالهای آبي رنگ نامحلول انجام می شود. در این روش برخلاف ساير روشها مراحل شستشو و هاروست كردن سلول كه اغلب باعث از دست رفتن تعدادي از سلولها می شوند حذف شده اند و تمام مراحل آزمايش از ابتداي كشت سلولي تا قرائت نتايج با فتومتر، در يك ميكروپليت انجام می شوند لذا تکرار پذیری، دقت و حساسیت آزمایش بالا است. بطور کلی سلولها در محیط کشت به دو صورت چسبیده به پلیت (Attached) و شناور (suspended) دیده می شوند. در صورتی که آزمایش روی سلولهای چسبیده به پلیت نظیر سلولهای توموری و فیبروبلاستها انجام شود ابتدا بایستی تعداد مناسبی سلول (ترجیحا 5000 سلول در هر چاهک) را در هر یک از چاهکها کشت داده اجازه داد تا سلولها به کف پلیت چسبیده و بحالت پایدار خود درآیند. سپس چاهکهای کنترل و آزمایش انتخاب شده و مقدار مناسبی از ماده میتوژن یا داروی مورد نظر را به چاهکهای تست اضافه می کنیم و پلیت را تا زمان مورد نیاز جهت تاثیر ماده مورد نظر، انکوبه می کنیم. پس از اتمام زمان انکوباسیون محيط كشت روئي را دور ريخته به هر چاهك 200 ميكروليتر محيط كشت حاوي نيم ميليگرم در ميلي ليتر محلول MTT اضافه نموده و به مدت 2 تا 4 ساعت در انكوباتورCO2 در 37 درجه سانتيگراد قرار می دهیم. در طی زمان انکوباسیون MTT توسط سيستم سوكسينات دهیدروژناز كه يكي از آنزيمهاي چرخه تنفسي ميتوكندريهاست احیا می شود. احیا و شکسته شدن این حلقه موجب تولید کریستالهای آبی رنگ فورمازان می شود که در زیر میکروسکوپ براحتی قابل تشخیص هستند. ميزان رنگ توليد شده با تعداد سلولهايي كه از نظر متابوليك فعال هستند رابطه مستقيم دارد. کریستالهای فورمازان در آب غيرمحلول بوده و بايستي قبل از رنگ سنجی توسط ماده حلالي نظير DMSO بحالت محلول درآيند. در نهایت جذب نوری محلول بدست آمده را می توان در طول موج 570 نانومتر قرائت کرده و به کمک منحنی استاندارد تعداد سلولها را محاسبه نمود. برای هر رده سلولی ارتباط خطی بین تعداد سلولها و جذب نوری محلول نهائی وجود دارد لذا جهت بررسی هر نوع سلول بایستی منحنی استاندارد مربوط به همان رده را رسم نموده و استفاده کرد.
انجام آزمایش روی سلولهاي معلق یک اختلاف جزئی با سلولهای چسبیده دارد. در مورد سلولهای هیبریدوما تعداد 5000 سلول در هر چاهک برای آزمایش مناسب است. اما در مورد لنفوسیتهای انسانی ده برابر سلول بیشتری لازم داریم و معمولا بتعداد 25000 تا 250000 سلول در هر چاهک رنگ نهائی مناسبی می دهد. در مورد سلولهای معلق باید پس از اتمام مراحل كار كشت سلولي پليت را در سانتريفيوژ مخصوص قرار داده و در دور 1000-2000 دور در دقيقه سانتريفيوژ نمود تا سلولهاي معلق در كف پليت رسوب كنند. در صورت لزوم می توان سلولها را با الكل يا فرمالدئيد در كف پليت فيكس نمود. عمل فيكساسيون بايستي در زمان كوتاهي انجام شود تا خصوصيات آنزيمي سلولها حفظ گردد. سپس سلولها را با PBS سرد شستشو داده و بر روي آنها محيط كشت حاوي MTT مي افزائيم. بقیه مراحل شبیه سلولهای چسبیده می باشد.
براي تهيه منحني استاندارد ابتدا يك سوسپانسيون سلولي با غلظت كاملاً مشخص مثلاً يك ميليون سلول در ميلي ليتر از سلولهاي مورد نظر تهيه كرده و پس از بررسي ويابيليتي آنها مقادير مشخص و فزاينده اي از آن را بصورت quadriuplicate در پليت 96 حفره اي كشت سلولي كشت مي دهيم. سپس اين پليت را به انكوباتور CO2 منتقل نموده و 24 ساعت بعد تست MTT انجام داده و با بدست آوردن مقادیر جذب نوری در مقابل تعداد سلول مربوطه منحني استاندارد را رسم می کنیم.
REFERENCES
1. Falak Reza ; (2001), M.S. Thesis. Tehran University, Iran.
2. Khorramizadeh M. R. et al.; Iranian J. Allergy, Asthma & Immunol. 2004; 3, 1, 7-11.
3. Khorramizadeh et al. (2002) Poster presen. 20th World Cong. of derm. PARIS.
4. Mosmann T. (1983) J. Immunol. Methods 65, 55–63.
5. Tada H. et al. (1986) J. Immunol. Methods 93, 157–165.
6. Denizot F. et al. (1986) J. Immunol. Methods 89, 271–277.
7. Gerlier, D. et al. (1986) J. Immunol. Methods 94, 57–63.
8. Hansen, M. B. et al. (1989) J. Immunol. Methods 119, 203–210.
9. Heeg K. et al. (1985) J. Immunol. Methods 77, 237–246.
10. Mosmann, T. et al. (1989) J. Immunol. Methods 116, 151–158.
11. Ohno M. et al. (1991) J. Immunol. Methods 145, 199–203.
12. Berg K. et al. (1988) J. Interferon Res. 8, (suppl. 1), S. 67.
13. Green, L. M. et al. (1984) J. Immunol. Methods 70, 257–268.
14. Espevik T. et al. (1986) J. Immunol. Methods 95, 99–105.
15. Ferrari M. et al. (1990) J. Immunol. Methods 131, 165–172.
16. Van de Loosdrecht et al. (1991) J. Immunol. Methods 141, 15–22.
17. Berridge M. V. et al. (1993) Arch. Biochem. Biophys. 303, 474-482.
18. Altman F. P (1976) Prog. Histochem. Cytochem. 9(3):1-56.
19. Kirkpatrick D. L. et al. (1990) Leukemia. Res. 14:459-466.
20. Liu Y. B. et al. (1997); J. Neurochem. 69:581-593.
21. Roehm N. W. et al. (1991) J. Immunol. Methods 142:257-265.
22. Sieuwerts A. M. et al. (1995) Eur. J. Clin. Chem. Clin. Biochem. 33:813-823.
پروتئينهاي شوك حرارتي به مجموع پروتئينهايي گفته مي شوند كه در شرايط استرس در سلول بيان مي گردند. نقش اين ملكولها جلوگيري از تغيير ساختار پروتئينها تحت عوامل استرسي مي باشد. اين پروتئينها در همه سلولهاي زنده در وضعيت متصل يا غير متصل به پروتئينها وجود دارند. مكان اين پروتئينها در هسته و سيتوپلاسم سلول مي باشد پروتئينهاي. HSP بعنوان چاپرونهاي مولكولي در فرايند هاي متعددي همچون فولدينگ پروتئينها، تجمع و انتقال آنها، عبور و مرور پپتيد ها و پردازش آنتي ژن تحت شرايط فيزيولوژيك و استرسي نقش دارند. بيان پروتئينهاي HSP بواسطه چندين نوع از عوامل استرس زا همچون تب، الكل، التهاب، استرسهاي اكسيداتيو، فلزات سنگين و همچنين شرايطي كه موجب جراحت و نكروزيس مي گردد، القاء مي شوند. هنگام افزايش سنتز پروتئين(مثل عفونتهاي ويروسي) و استرس، پروتئينهاي HSPبه جايگاه هاي هيدروفوبيك روي پلي پپتيد ها متصل و سبب بروز تغييرات ساختاري در آنها مي گردند. از سوي ديگر، سبب جلوگيري از ايجاد پروتئينهايي با تا خوردگي يا فولدينگ اشتباه مي شوند. ضمناً، از واكنش نامناسب آنها با ملكولهاي ديگر جلوگيري مي كنند. همچنينHSP ،كمك به انتقال ملكولها در عرض غشاء، ، تشكيل كمپلكس هاي مولتي مر( (Multimeric Complexes، و تغيير فعاليت هاي سوبسترائي ملكولهاي ناشناس (مانند پلي پپتيدهاي ويروسي) مي كند. نسخه برداري از ژنهاي HSP از طريق تعامل بين فاكتورهاي رونويسي يا HSF((Heat shock factorsبا عناصر شوك حرارتي يا HSE (Heat Shock Eelements)در نواحي پروموتور ژنهاي HSP صورت مي گيرد .
استفاده از اين ملكولها جهت ايمني درماني تومور مزيتهائي دارد زيرا، اين ملكولها بعنوان سيستم هاي حمل طبيعي پپتيدها با چندين عملكرد شناخته شده اند. اولاً، HSP قادرند به پپتيدهاي آنتي ژنيك متصل و آنها را به سلولهاي عرضه كننده آنتي ژن حمل كنند. ثانياً، HSP به راحتي توسط سلولهاي عرضه كننده آنتي ژن از طريق رسپتورهاي اندوسيتيك برداشته مي شوند. ثالثاً ، HSP خود بعنوان ادجوانت عمل مي كند و باعث فعال شدن سلولهاي ايمني ذاتي و توليد سيتوكينهاي التهابي مي شود. ماهيت و چگونگي عرضه كمپلكس هاي-HSP پپتيد ناشناخته است.مطالعات نشان داده اند كه، پس از اندوسيوز كمپلكس
-HSP پپتيد، دو مسير متفاوت در عرضه پپتيد بر روي MHC I وجود دارد. اين دو مسير شامل مسير وابسته به TAP(Transporter-associated with Antigen Processing)و پروتئازوم سيتوزولي و يا مسير اندوزومي مستقل از TAP و پروتئازوم مي باشد.
پس از اندوسيتوز، قسمتي از كمپلكس هاي-HSP پپتيد از طريق مكانيسمهاي ناشناخته اي به سيتوزول منتقل و فرايندهاي سيتوزولي بر روي آنها انجام مي شود. در مكانيسم وابسته به مسير اندوزومي، فرايندهاي اندوزومي بر روي كمپلكس هاي-HSP پپتيد انجام و در انتها با مكانيسم ناشناخته اي از اندوزوم/فاگوزوم خارج، به سيتوزول داخل و سپس بر روي MHC Iعرضه مي شود. اين فرايند تحت عنوان عرضه متقاطع بر روي MHC I و تحريك متقاطع شناخته شده است و اولين بار توسط Srivastava و همكاران كشف شد. البته مكانيسمهاي نا شناخته ديگري نيز ممكن است در عرضه آنتي ژنهاي با منشاء خارجي بر روي MHC I دخيل باشند
پروتئينهاي شوك حرارتي بر حسب وزن ملكولي شان به سه خانواده عمده شامل 50،60 ، 70 و 90 كيلو دالتوني تقسيم مي شوند. همچنين مي توان آنها را بر حسب محل داخل سلولي به زيرگروه هاي ديگري تقسيم كرد. مثلاً پروتئينهاي مرتبط با گلوكز(GRPs) زيرگروهي هستند كه مقيم شبكه اندوپلاسميك مي باشند. اهميت ايمنولوژيك HSP بر اساس خاصيت ادجوانتي و قابليت آنها در عرضه متقاطع آنتي ژنها بر روي MHC I مي باشد كه آنها را بعنوان واكسنهاي كانديد در ايمني
درماني تومور مطرح كرده است. از جمله اعضاي اين خانواده كه خواص ضد توموري داشته اندمي توان ازHSP110 ،HSP90 ،HSP 70 (از اعضاي خانواده 70 (HSP، GRP170 ،GP96 و كالرتيكولين نام برد. به دليل خواص ادجوانتي و توليد سيتوكينهاي التهابي، از اين ملكولها تحت عناوين ايمنوادجوانت و يا چاپروكاين نيز ياد شده است. در يك مطالعه، قدرت ايمني زائي ضد تومور بين HSP70 ، HSP90 و 96 GP مقايسه و در آن نشان داده شد كه خاصيت ضد توموري HSP70 و GP 96 يكسان و بيش از 90 HSP مي باشد. قبلاً، واكسن پلاسميدي حاوي HSP70 و Her 2 بشكل فيوژن و يا واكسن پروتئيني حاوي HSP 110 و Her 2 نوتركيب ، كه در شرايط آزمايشگاه بهم متصل شده بودند، بر روي يك مدل تومور Her 2 مثبت بررسي شده است. اثرات افزايشي GP 96 بر ايمني درماني با استفاده از آنتي ژن Her 2 تاكنون مطالعه نشده است. با توجه به قابليت هاي GP 96 در تحت تأثير قراردادن سيستم ايمني با استفاده از آنتي ژن عوامل عفوني نشان داده شده است، انتظار مي رود GP96 همراه Her2 قادر به ايجاد يك پاسخ ايمني مناسب و قوي باشد. به اين منظور نياز به مطالعه بيشتر جزئيات در خصوص ساختمان، رفتار بيولوژيك و ايمنولوژيك GP96 مي باشد.
